第三节 肾小管与集合管的转运功能
人两肾每天生成的肾小球滤过液达180L,而终尿仅为1.5L。这表明滤过液中约99%的水被肾小管和集合管重吸收,只有约1%被排出体外。不仅如此,滤过液中的葡萄糖已全部被肾小管重吸收回血;钠、尿素告示不同程度地重吸收;肌酐、尿酸和K[SB]+[/SB]等还被肾小管分泌入管腔中。
一、肾小管与集合管的转运方式
肾小管和集合管的转运包括重吸收和分泌。重吸收是指物质从肾小管液中转运至血液中,而分泌是指上皮细胞本身产生的物质或血液中的物质转运至肾小管腔内。肾不球滤过液进入肾小管后称为小管液。
物质通过细胞的转运包括被动转运和主动转运。被动转运是指溶质顺电化学梯度通过肾小管上皮细胞的过程。水的渗透夺之差是水的转运动力。水从渗透压低一侧通过细胞膜进入渗透压高一侧。
主动转运是指溶质逆电化学梯度通过肾小管上皮细胞的过程。主动转运需要消耗能量,根据主动转运过程中能量来源的不同,分为原发性主动转运和继发性主动转运。原发性主动转运(简称为主动转运)所需要消耗的能量由ATP水解直接提供。例如Na[SB]+[/SB]和K[SB]+[/SB]的主动转运是靠细胞膜上的Na[SB]+[/SB]泵水解ATP直接提供能量的。继发性主动转运所需的能量不是直接来自Na[SB]+[/SB]泵。而是来自其他溶质顺电化学梯度转运时释放的。例如一些物质的继发性主动转运的动力直接来自Na[SB]+[/SB]顺电化学梯度转运时释放的能量。释放的之些能量归根到底也是来自Na[SB]+[/SB]泵。由于上皮细胞基侧膜上存在Na[SB]+[/SB],将细胞内的Na[SB]+[/SB]泵至细胞外,造成细胞内的Na[SB]+[/SB]浓度明显低于细胞外,细胞外K[SB]+[/SB]被泵回细胞内,造成细胞内K[SB]+[/SB]浓度明显高于细胞外,并维持细胞内的负电位。这样,小管液中的Na[SB]+[/SB]便顺电化学梯度通过管腔膜进入细胞,并释放能量提供其他物质的转运。许多物质的转运都与Na[SB]+[/SB]的主动转运相耦联,例如小管液中的葡萄糖、氨基酸、有机酸和CI[SB]-[/SB]等物质的重吸收都与Na[SB]+[/SB]同向转运(cotransport)有关。同向转运是指两种物质与细胞膜上的同向转运体(cotransporter,symporter)特殊蛋白质结合,以相同方向通过细胞膜的转运;又如肾小管细胞分泌H[SB]+[/SB]是与Na[SB]+[/SB]的逆向转运相耦联。逆向转运(antiport)是指两种物质与细胞膜上的逆向转运体(antiport)又称交换体(exchanger)结合,以相反方向通过细胞膜的转运。可见,Na[SB]+[/SB]的主动转运在肾小管上皮细胞的转运中起着关键作用(图8-9)。一个带下电荷和另一个带负电荷的两种物质的同向转动,或电荷相同的两种物质的逆向转运都不会造成小管内外电位改变,这种转运称为电中性转动。如果一个物质是离子,另一个是电中性物质,这种转运就会使小管内外出现电位差,称为生电性转运。如在近球小管,Na[SB]+[/SB]与葡萄糖的同向转运,因葡萄糖是电中性物质,Na[SB]+[/SB]和葡萄糖被重吸收就会造成小管内较小管外带负电位。又如在近球小管的后半段,小管液CI[SB]-[/SB]浓度比管外高,CI[SB]-[/SB]顺浓度差被动重吸收造成管内带正电位。
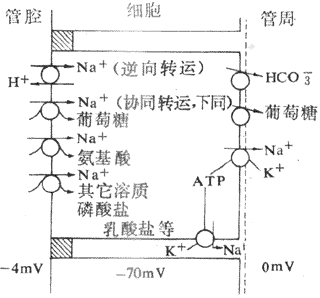
图8-9 Na[SB]+[/SB]转运与其他溶质转运之间的伴联关系
二、各段肾小管和集合管的转运功能
(一)近球小管
肾小球滤过流经近球小管后,滤过液中67%Na[SB]+[/SB]、CI[SB]-[/SB]、K[SB]+[/SB]和水被重吸收,85%的HCO[XB]3[/XB]也被重吸收,葡萄糖、氨基酸全部被重吸收;H[SB]+[/SB]则分泌到肾小管中。近球小管重吸收的关键动力是基侧膜上的Na[SB]+[/SB]泵;许多溶质,包括水的重吸收都与Na[SB]+[/SB]泵的活动有关。
1.Na[SB]+[/SB]、CI[SB]-[/SB]和水的重吸收在近球小管前半段,大部分Na[SB]+[/SB]与葡萄糖,氨基酸同向转运、与H[SB]+[/SB]逆向转运而被主动重吸收;面近球小管后半段,Na[SB]+[/SB]和CI[SB]-[/SB]主要通过细胞旁路而被被动重吸收。水随NaCI等溶质重吸收而被重吸收,因此,该段小管注与血浆渗透压相同,是等渗重吸收。
在近球小管前半段,由于Na[SB]+[/SB]泵的作用,Na[SB]+[/SB]被泵至细胞间隙,使细胞内Na[SB]+[/SB]浓度低,细胞内带负电位。因此,小管液中的Na[SB]+[/SB]和葡萄糖与管腔膜上的同向转运体结合后,Na[SB]+[/SB]顺电化学梯度通过管腔膜的同时,释放的能量将葡萄糖同向转运入细胞内。进入细胞内的Na[SB]+[/SB]即被细胞基侧膜上的Na[SB]+[/SB]泵泵出至细胞间隙,这样,一方面使细胞内Na[SB]+[/SB]的浓度降低,小管液中的Na[SB]+[/SB]-葡萄糖便可不断转运进入细胞内,细胞内的葡萄糖由易化扩散通过细胞基侧膜离开细胞回到血液中;另一方面,使细胞间隙中的Na[SB]+[/SB]浓度升高,渗透压也升高,通过渗透作用,水随之进入细胞间隙。由于细胞间隙在管腔膜侧的紧密连接相对是密闭的,Na[SB]+[/SB]和水进入后就使其中的静水压升高,这一压力可促使Na[SB]+[/SB]和水通过基膜进入相邻的毛细血管而被重吸收,但也可能使部分Na[SB]+[/SB]和水通过紧密连接回漏(back-leak)至小管腔内(图8-10A)。
另一部分的Na[SB]+[/SB]-H[SB]+[/SB]交换而主动重吸收。小管液中的Na[SB]+[/SB]和细胞内的H[SB]+[/SB]与管腔膜上的交换体结合进行逆向转运,使小管液中的Na顺浓度梯度通过管腔膜进入细胞的同时,将细胞内的H[SB]+[/SB]分泌到小管液中;进入细胞内的Na[SB]+[/SB]随即被基侧膜上的Na[SB]+[/SB]泵泵至细胞间隙而主动重吸收。分泌到小管液中的H[SB]+[/SB]将有利于小管液中的HCO[XB]3[/XB]的重吸收。
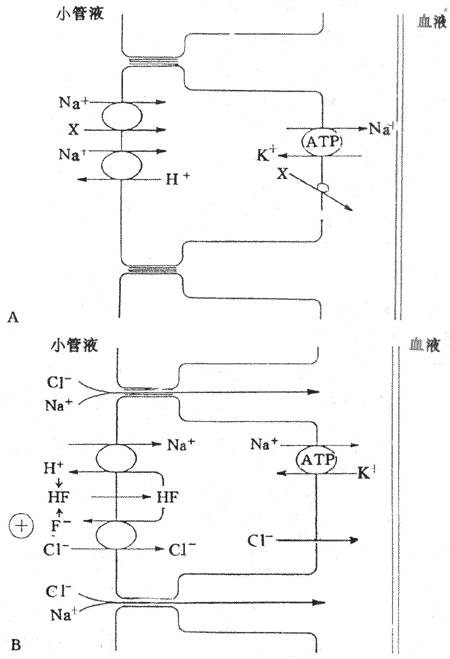
图8-10 近球小管重吸收NaCI的示意图
A:近球小管的前半段 X代表葡萄糖、氨基酸、磷酸盐CI[SB]-[/SB]
B:近球小管的后半段F[SB]-[/SB]代表甲酸盐 HF,甲酸
在近球小管后半段,NaCI是通过细胞旁路和跨上皮细胞两条途径而被重吸收的。小管液进入近球小管后半段时,绝大多数的葡萄糖、氨基酸已被重吸收。由于HCO[XB]3[/XB]重吸收速率明显大于CI[SB]-[/SB]重吸收,CI[SB]-[/SB]留在小管液中,造成近球小管后半段的CI[SB]-[/SB]浓度比管周组织间液高20%-40%。因此,CI[SB]-[/SB]顺浓度梯度经细胞旁路(即通过紧密连接进入细胞间隙)而重吸收回血。由于CI[SB]-[/SB]被动重吸收是生电性的,使小管液中正离子相对较多,造成管内外电位差,管腔内带正电,管外带负电,在这种电位差作用下,Na[SB]+[/SB]顺电位差通过细胞旁路而被动重吸收。CI[SB]-[/SB]通过细胞旁路重吸收是顺浓度梯度进行的,而Na[SB]+[/SB]通过细胞旁路重吸收是顺电位梯度进行的,因此,NaCI是重吸收都是被动的(图8-10B)。
NaCI跨上皮细胞重吸收与H[SB]+[/SB]和甲酸盐(formate,F[SB]-[/SB])再循环有关,要通过管腔膜上相互耦联的Na[SB]+[/SB]-H[SB]+[/SB]交换和CI[SB]—[/SB]甲酸根交换。在正常肾小管液中含有低浓度甲酸根,通过Na[SB]+[/SB]、H[SB]+[/SB]交换,Na[SB]+[/SB]进入细胞,H[SB]+[/SB]分泌到小管液中,并与小管液中的甲酸根结合形成甲酸(formic acid,HF)。甲酸是脂溶性的,可迅速通过管腔膜进入细胞,在细胞内分解为H[SB]+[/SB]和甲酸根。甲酸根和小管液中的CI[SB]-[/SB]进行逆向转运,结果,CI[SB]-[/SB]进入细胞内并通过基侧膜而被重吸收,而甲酸根则人细胞内进入小管液。细胞内的H[SB]+[/SB]则与小管液中的Na[SB]+[/SB]进行逆向交换,Na[SB]+[/SB]进入细胞,并被Na[SB]+[/SB]泵泵至细胞间隙,然后进入管周毛细血管而被重吸收;H[SB]+[/SB]分泌至小管液,再与小管液中的甲酸根据结合,形成甲酸再进入细胞。因此,H[SB]+[/SB]和甲酸根可再循环使用,CI[SB]-[/SB]和Na[SB]+[/SB]则被重吸收回血。
水的重吸收是被动的,是靠渗透作用而进行的。水重吸收的渗透梯度存在于上皮细胞和细胞间隙之间。这是由于Na[SB]+[/SB]顺电化学梯度通过管腔膜进入细胞后,细胞内的Na[SB]+[/SB]被基侧膜上的Na[SB]+[/SB]泵泵至细胞间隙,使细胞间隙渗透压升高。在渗透作用下,水便不断从小管液进入上皮细胞,并从细胞不断进入细胞间隙,造成细胞间隙静水压升高;加上管周毛细血管内静水压较低,胶体渗透压较高,水便通过周围组织间隙进入毛细血管而被重吸收。
2.HCO[XB]3[/XB]重吸收与H[SB]+[/SB]的分泌 HCO[XB]3[/XB]的重吸收与小管上皮细胞管腔膜上的Na[SB]+[/SB]-H[SB]+[/SB]交换有密切关系。HCO[XB]3[/XB]在血浆中钠盐(NaHCO[XB]3[/XB])的形式存在,滤过中的NaHCO[XB]3[/XB]滤入囊腔进入肾小管后可解离成Na[SB]+[/SB]和HCO[XB]3[/XB]。通过Na[SB]+[/SB]-H[SB]+[/SB]交换,H[SB]+[/SB]由细胞内分泌到小管液中,Na[SB]+[/SB]进入细胞内,并与细胞内的HCO[XB]3[/XB]一起被转运回血(图8-11)。由于小管液中的HCO[XB]3[/XB]不易通过管腔膜,它与分泌的H[SB]+[/SB]结合生成H[XB]2[/XB]CO[XB]2[/XB],在碳酸酐酶作用下,H[XB]2[/XB]CO[XB]2[/XB]迅速分解为CO[XB]2[/XB]和水。CO[XB]2[/XB]是高度脂溶性物质,能迅速通过管腔膜进入细胞内,在碳酸酐酶作用下,进入细胞内的CO[XB]2[/XB]与H[XB]2[/XB]O结合生成H[XB]2[/XB]CO[XB]3[/XB]。H[XB]2[/XB]CO[XB]3[/XB]又解离成H[SB]+[/SB]和HCO[XB]3[/XB]。H[SB]+[/SB]通过Na[SB]+[/SB]-H[SB]+[/SB]交换从细胞分泌到小管液中,HCO[XB]3[/XB]则与Na[SB]+[/SB]一起转运回血。因此,肾小管重吸收HCO[XB]3[/XB]是以CO[XB]2[/XB]的形式,而不是直接以HCO[XB]3[/XB]的形式进行的。如果滤过的HCO[XB]3[/XB]超过了分泌的H[SB]+[/SB],HCO[XB]3[/XB]就不能全部(以CO[XB]2[/XB]形式)被重吸收。由于它不易透过管腔膜,所以余下的便随尿排出体外。可见肾小管上皮细胞分泌1H[SB]+[/SB]就可使1HCO[XB]3[/XB]和1Na[SB]+[/SB]重吸收回血,这在体内的酸碱平衡调节中起到重要作用。乙酰唑胺可抑制碳酸酐酶的活性,因此,用乙酰唑受后,Na[SB]+[/SB]-H[SB]+[/SB]交换就会减少,因而NaHCO[XB]3[/XB]、NaCI和水的排出增加,可引起利尿。由于近球小管的Na[SB]+[/SB]-H[SB]+[/SB]交换,小管液中的HCO[XB]3[/XB]与H[SB]+[/SB]结合并生成CO[XB]2[/XB],CO[XB]2[/XB]透过管腔膜的速度明显高于CI[SB]-[/SB]的速度。因此,HCO[XB]3[/XB]的重吸收率明显大于CI[SB]-[/SB]的重吸收率。
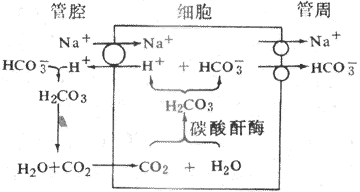
图8-11 肾小管上皮细胞生成和分泌H[SB]+[/SB]示意图
3.K[SB]+[/SB]的重吸收微穿刺实验表明,肾小球滤过的K[SB]+[/SB],67%左右在近球小管重吸收回血,而尿中的K[SB]+[/SB]主要是由远曲小管和集合管分泌的。有人认为,近球小管对K[SB]+[/SB]的重吸收是一个主动转运过程。小管液中钾浓度为4mmol/L,大大低于细胞内K[SB]+[/SB]浓度(150mmol/L)。因此在管腔膜处K[SB]+[/SB]重吸收是逆浓度梯度进行的。管腔膜K[SB]+[/SB]主动重吸收的机制尚不清楚。
4.葡萄糖重吸收肾小球滤过液中的葡萄糖浓度与血糖浓度相同,但尿中几乎不含葡萄糖,这说明葡萄糖全部被重吸收因血。微穿刺实验表明,重吸收葡萄糖的部位仅限于近球小管,尤其是在近球小管前半段,其他各段肾小管都没有重吸收葡萄糖的能力。因此,如果在近球小管以后的小管液中仍含有葡萄糖,则尿中将出现葡萄糖。
葡萄糖是不带电荷的物质,它的逆浓度梯度重吸收的,是由Na[SB]+[/SB]继发性主动同向转运而被重吸收的。在兔肾近球小管微灌流实验中观察到,如果灌流液中去掉葡萄糖等有机溶质,则Na[SB]+[/SB]的重吸收率降低;如果灌流液中的Na[SB]+[/SB]全部去掉,则葡萄糖有机溶质的重吸收将完全停止,说明葡萄糖的重吸收与Na[SB]+[/SB]同向转运密切相关。葡萄糖和Na[SB]+[/SB]分别与管腔膜上的同向转运体蛋白的结合位点相结合而进行同向转运(见前述Na[SB]+[/SB]重吸收)。
近球小管对葡萄糖的重吸收有一定限度。当血液中葡萄糖浓度超过160-180mg/100ml时,有一部分肾小管对葡萄糖的吸收已达到极限,尿中开始出现葡萄糖,此时的血糖浓度称为肾糖阈。血糖浓度再继续升高,尿中葡萄糖含量也将随之不断增加;当血糖浓度超过300mg/100ml后,全部肾小管对葡萄糖的吸收均已达到极限,此值即为葡萄糖吸收极限量。此时,尿葡萄糖排出率则随血糖浓度升高而平行增加。人肾的葡萄糖吸收极限量,在体表面积为1.73m[SB]2[/SB]的个体,男性为375mg/min,女性为300mg/min。肾之所以有葡萄糖吸收极限量,可能是由于同向转运体的数目有限的缘故,当所有同向转运体的结合位点都被结合而达饱和时,葡萄糖转运量就无从再增加了。
5.其他物质的重吸收和分泌小管注保的氨基酸的重吸收与葡萄糖的重吸收机制相同,也与Na[SB]+[/SB]同向转运(图8-9)。但是,转运葡萄糖的和转运氨基酸的同向转运体可能不同,也就是说同向转运体具有特异性。此外,HPO[XB]4[/XB][SB]-2[/SB]、SO[XB]4[/XB][SB]-2[/SB]的重吸收也也Na[SB]+[/SB]同向转运而进行。正常时进入滤液中的微量蛋白质则通过肾小管上皮细胞吞饮作用而被重吸收。
体内代谢产物和进入体仙的某些物质如青霉素、酚红,大部分的利尿药等,由于与血浆中蛋白结合而不能通过肾小球滤过,它们均在近球小管被主动分泌到小管液中而排出体外。
(二)髓袢
近球小管液流经髓袢过程中,约20%的Na[SB]+[/SB]、CI[SB]-[/SB]、和K[SB]+[/SB]等物质被进一步重吸收。髓袢升支粗段的NaCI重吸收在尿液稀释和浓缩机制中具有重要意义。髓袢升支粗段CI[SB]-[/SB]是逆电化学梯度被上皮细胞重吸收的。微穿刺实验证明,兔髓袢升支粗段管腔内为正电位(+10mV)。在微灌流实验中,如果灌流液中不含K[SB]+[/SB],则管内的正电位基本消失,CI[SB]-[/SB]重吸收率很低,这说明管腔内正电位与CI[SB]-[/SB]的重吸收和小管液中的K[SB]+[/SB]有密切关系。如果在髓袢升支粗段管周的浸溶液中加入选择性Na[SB]+[/SB]泵抑制剂哇巴因(ouabain)抑制Na[SB]+[/SB]泵后,CI[SB]-[/SB]的转运也受阻,说明Na[SB]+[/SB]泵是CI[SB]-[/SB]重吸收的重要因素。据上述实验,有人提出Na[SB]+[/SB]:2CI[SB]-[/SB]:K[SB]+[/SB]同向转运模式来解释升支NaCI的继发性主动重吸收。该模式认为:①髓袢升支粗段上皮细胞基侧膜上的Na[SB]+[/SB]泵,将Na[SB]+[/SB]由细胞内示向组织间液,使细胞内的Na[SB]+[/SB]下降,造成管腔内与细胞内Na[SB]+[/SB]有明显的浓度梯度;②Na[SB]+[/SB]与管腔膜上同向转运体结合,形成Na[SB]+[/SB]:2CI[SB]-[/SB]:K[SB]+[/SB]同向转运体复合物,Na[SB]+[/SB]顺电化学梯度将2CI[SB]-[/SB]和K[SB]+[/SB]一起同向转运至细胞;③进入细胞内的Na[SB]+[/SB]、2CI[SB]-[/SB]和K[SB]+[/SB]的去向各不相同:Na[SB]+[/SB]由Na[SB]+[/SB]泵泵至组织间液,2CI[SB]-[/SB]由于浓度梯度经管周膜上CI[SB]-[/SB]通道进入组织间液,而K[SB]+[/SB]则顺浓度梯度经管腔膜而返回管腔内,再与同向转运体结合,继续参与Na[SB]+[/SB]:2CI[SB]-[/SB]K[SB]+[/SB]的同向转运,循环使用;④由于2CI[SB]-[/SB]进入组织间液,K[SB]+[/SB]返回管腔内,这就导致管腔内出现正电位;⑤由于管腔内正电位,使管腔液中的Na[SB]+[/SB]等正离子顺电位差从细胞旁路进入组织间液,这是不耗能的Na[SB]+[/SB]被动重吸收。从这个模式说明,通过Na[SB]+[/SB]泵的活动,继发性主动重吸收了2CI[SB]-[/SB],同时伴有2Na[SB]+[/SB]的重吸收,其中1Na[SB]+[/SB]是主动重吸收,另1Na[SB]+[/SB]通过细胞旁路而被动重吸收,这样为Na[SB]+[/SB]的重吸收节约了50%能量消耗(图8-12)。髓袢升支粗段对水的通透性很低,水不被重吸收而留在小管内。由于NaCI被上皮细胞重吸收至组织间液,因此造成小管液低渗,组织间液高渗。这种水和盐重吸收的分离,有利于尿液的浓缩和稀释。Na[SB]+[/SB]:2CI[SB]-[/SB]:K[SB]+[/SB]同向转运对速尿,利尿酸等利尿剂很敏感。它们与同向转运体结合后,可抑制其转运功能,管腔内正电位消失,NaCI的重吸收受抑制,从而干扰尿的浓缩机制,导致利尿。
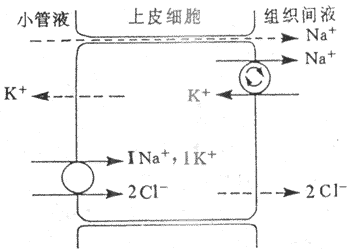
图8-12 髓袢升支粗段继发性主动吸收CI[SB]-[/SB]的示意图
(三)远球小管和集合管
在远曲小管和集合管,重吸收大约12%滤过的Na[SB]+[/SB]和CI[SB]-[/SB],分泌不同量的K[SB]+[/SB]和H[SB]+[/SB],重吸收不同量的水。水、NaCI的重吸收以及K[SB]+[/SB]和H[SB]+[/SB]的分泌可根据机体贴的水、盐平衡状况来进行调节。如相机缺水或缺盐时,远曲小管和集合管可增加水、盐的重吸收;当机体水、盐过剩时,则水、盐重吸收明显减少,水和盐从尿排出增加。因此,远曲小管和集合管对水和盐的转运是可被调节的。水的重吸收主要受抗利尿激素调节,而Na[SB]+[/SB]和K[SB]+[/SB]的转运主要受醛固酮调节。
远曲小管和集合管上皮细胞间隙的紧密连接对小离子如Na[SB]+[/SB]、K[SB]+[/SB]和CI[SB]-[/SB]等的通透性低,这些离子不易通过紧密连接回漏至小管腔内,因此,所能建立起来的管内外离子浓度梯度和电位梯度大。在远曲小管初段,对水的通透性很低,但仍主动重吸收NaCI,继续产生低渗小管液。Na[SB]+[/SB]在远曲小和集合管的重吸收是逆较大的电化学梯度进行的,是主动重吸收过程。这可能与远曲小管的Na[SB]+[/SB]泵在肾单位中活性最高有关。有人认为在远曲小管初段的小管液中,Na[SB]+[/SB]是通过Na[SB]+[/SB]-CI[SB]-[/SB]同向转运进入细胞的,然后由Na[SB]+[/SB]泵将Na[SB]+[/SB]泵出细胞而主动重吸收回血(图8-13A)。NaCI同向转运体可被噻嗪类利尿药所抑制。
远曲小管后段和集合管含有两类细胞,即主细胞和闰细胞。主细胞重吸收Na[SB]+[/SB]和水,分泌K[SB]+[/SB],闰细胞则主要分泌H[SB]+[/SB],主细胞重吸收Na[SB]+[/SB]主要通过管腔膜上的Na[SB]+[/SB]通道。管腔内的Na[SB]+[/SB]顺电化学梯度通过管膜上的Na[SB]+[/SB]通道进入细胞,然后,由Na[SB]+[/SB]泵泵至细胞间液而被重吸收(图8-13B)。
K[SB]+[/SB]的分泌 尿中K[SB]+[/SB]的排泄量视K[SB]+[/SB]的摄入量而定,高钾饮食可排出大量的钾,低钾饮食则尿中排钾量少,使机体的钾摄入量与排出量保持平衡,维持机体K[SB]+[/SB]浓度的相对恒定。
K[SB]+[/SB]分泌的动力包括:①在远曲小管和集合管的小管液中,Na[SB]+[/SB]通过主细胞的管腔膜上的Na[SB]+[/SB]通道进入细胞,然后,由基侧膜上的Na[SB]+[/SB]泵将细胞内的Na[SB]+[/SB]泵至细胞间隙而被重吸收,因而是生电性的,使管腔内带负电位(-10?40mV)。这种电位梯度也成为K[SB]+[/SB]从细胞分泌至管腔的动力;②在远曲小管后段和集合管的主细胞内的K[SB]+[/SB]浓度明显高于小管液中的K[SB]+[/SB]浓度,K[SB]+[/SB]便顺浓度梯度从细胞内通过管腔膜上的K[SB]+[/SB]通道进入小管液;③Na[SB]+[/SB]进入主细胞后,可刺激基侧膜上的Na[SB]+[/SB],使更多的K[SB]+[/SB]从细胞外液中泵入细胞内,提高细胞内K[SB]+[/SB]浓度,增加细胞内和小管液之间的K[SB]+[/SB]浓度梯度,从而促进K[SB]+[/SB]分泌,因此,K[SB]+[/SB]的分泌与Na[SB]+[/SB]的重吸收有密切关系。(图8-13B)。
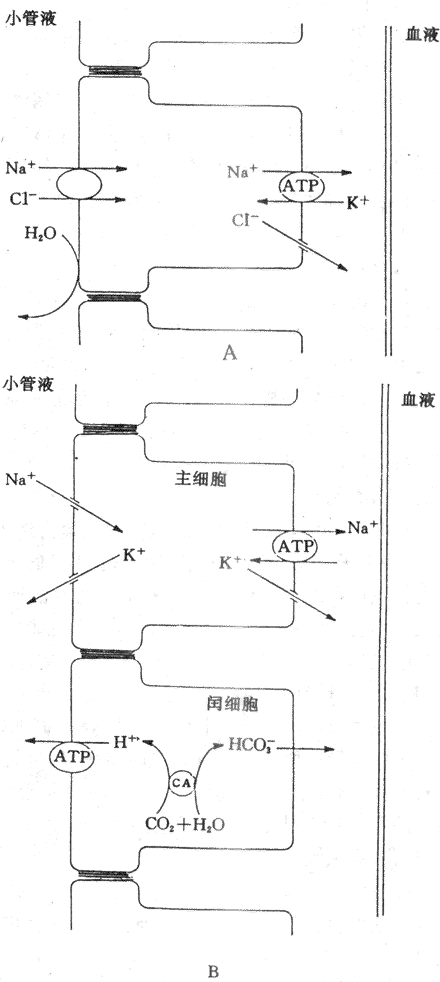
图8-13 远球小管和集合管重吸收NaCI、分泌K[SB]+[/SB]和H[SB]+[/SB]的示意图
A:远曲小管初段 B:远曲小管后段和集合管
H[SB]+[/SB]的分泌除了近球小管细胞通过Na[SB]+[/SB]-H[SB]+[/SB]交换分泌H[SB]+[/SB],促进NaHCO[XB]3[/XB]重吸收外,远曲小管和集合管的闰细胞也可分泌H[SB]+[/SB]。H[SB]+[/SB]的分泌是一个逆电化学梯度进行的主动转运过程。有人认为管腔膜上有H[SB]+[/SB]泵,能将细胞内的H[SB]+[/SB]历史意义入小管腔内。细胞内的CO[XB]2[/XB]和H[XB]2[/XB]O在碳酸酐酶催化作用下生成的H[SB]+[/SB]和HCO[XB]3[/XB],H[SB]+[/SB]由H[SB]+[/SB]泵泵至小管液,HCO[XB]3[/XB]则通过基侧膜回到血液中,因而H[SB]+[/SB]分泌和HCO[XB]3[/XB]的重吸收与酸碱平衡的调节有关(图8-13B)。闰细胞分泌的H[SB]+[/SB]与HPO[XB]4[/XB][SB]-2[/SB]结合形成H[XB]2[/XB]PO[XB]4[/XB],这是可滴定酸;分泌的H[SB]+[/SB]可与上皮细胞分泌的NH[XB]3[/XB]结合,形成NH[XB]4[/XB][SB]+[/SB]。可滴定酸和NH[XB]4[/XB][SB]+[/SB]都不易透过管腔膜进入细胞而留在小管液中。因此,它们是尿液酸碱度的决定因素。
NH[XB]3[/XB]的分泌远曲小管和集合管的上皮细胞在代谢过程中不断地生成NH[XB]3[/XB],这些NH[XB]3[/XB]主要由谷氨酰胺脱氨而来。NH[XB]3[/XB]具有脂溶性,能通过细胞膜向小管液周围组织间液和小管液自由扩散,扩散量取决于两种液体的pH值。小管液的pH较低(H[SB]+[/SB]浓度较高),所以NH[XB]3[/XB]较易向小管液中扩散。分泌的NH[XB]3[/XB]能与小管液中的H[SB]+[/SB]结合并生成NH[XB]4[/XB][SB]-[/SB],小管液中NH[XB]3[/XB]浓度因而下降,于是管腔膜两侧形成了NH[XB]3[/XB]浓度梯度,此浓度梯度又加速了NH[XB]3[/XB]向小管液中扩散。由此可见,NH[XB]3[/XB]的分泌与H[SB]+[/SB]的分泌密切相关;H[SB]+[/SB]分泌增加促使NH[XB]3[/XB]分泌增多。NH[XB]3[/XB]与H[SB]+[/SB]结合并生成NH[XB]4[/XB][SB]-[/SB]后,可进一步与小管液中的强酸盐(如NaCI等)的负离子结合,生成酸性铵盐(NH[XB]4[/XB]CI等)并随尿排出。强酸盐的正离子(如Na[SB]+[/SB])则与H[SB]+[/SB]交换而进入肾小管细胞,然后和细胞内HCO[XB]3[/XB][SB]-[/SB]一起被转运回血。所以,肾小管细胞分泌NH[XB]3[/XB],不仅由于铵盐形成而促进了排H[SB]+[/SB],而且也促进了NaHCO[XB]3[/XB]的重吸收。