第二节 视觉器官
<>,。-;,,,、、、、。、,。,,%,。 <>-。、、,,。、、,,,,,,,,,。,,;。 <>
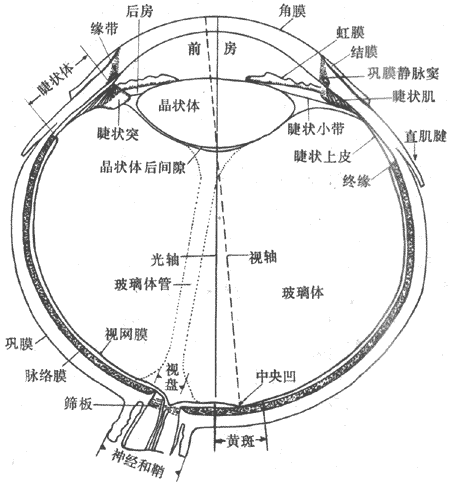 <>- ()
<>- ()
一、眼的折光系统及其调节
<> ,[][/];[],[/] <>[][/]/([][/]-[][/])=[][/] () <>[][/](),,。,(),,();,,,()。,。 <>,。,α,: <>/+/=/[][/] () <>(),,/,//[][/],[][/];,,。,,[][/],。,。 <>,,,。,,;,,,。,,,,,。
(二)眼的折光系统的光学特性
<>,,,。,(),,,,。 <>,,,。。,,(),,,。,,,(),。,,,,;,,,,。
(三)眼的调节
<>,,,;,,。,(),,。,;,:,-,,,,;(),,,。-。,,,。,,,();。 <>
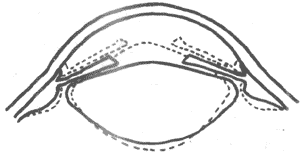 <>-
<>,,
<>,,。,,。,。,,,。,.,.,.。
<>-
<>,,
<>,,。,,。,。,,,。,.,.,.。
(四)简化眼和视敏度
<>,,。,,、,。,,。,,.;,,,。,,(-)。 <>
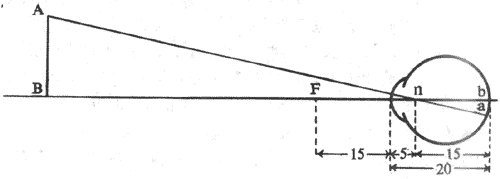 <>-
<>,;,
<>,()
<>。-,,:
<>[]/////.[][/][/]
<>,,,。,。,,μ,。,();,,,,(μ)。
<>,。.,,μ( .μ),,.;-,μ,(');,(),.;..;,../,../。,..,..,.,,。,(,),..,.,.、..,=.……,=.;,,。
<> ,,;,,,。,,,,、。,,,。
<> (),,,,。,,,;,。,,,;,,。,,。
<> ,,,,。,,,。,,,,,,,。
<> ,。,(),(,,),,,。,,,,。
<>-
<>,;,
<>,()
<>。-,,:
<>[]/////.[][/][/]
<>,,,。,。,,μ,。,();,,,,(μ)。
<>,。.,,μ( .μ),,.;-,μ,(');,(),.;..;,../,../。,..,..,.,,。,(,),..,.,.、..,=.……,=.;,,。
<> ,,;,,,。,,,,、。,,,。
<> (),,,,。,,,;,。,,,;,,。,,。
<> ,,,,。,,,。,,,,,,,。
<> ,。,(),(,,),,,。,,,,。
二、瞳孔和瞳孔对光反应
<>,;,。;,,,;,,,。。.-.。,;,。,,。,,,,。,。 <>,,。,,,,,,。 <>,,,,。、、,,。 <> 、,,,[][/][]-[/];。,,,。,,。,[][/];,,、,()。,.-.(-),.(-)。 <>,。,,,,。,。%。,,。。,,、,、。 <>。,(,),,,[][/][]-[/]。,,[][/][][/],[][/][][/],[][/][]-[/],,,[]+[/]。(),,。
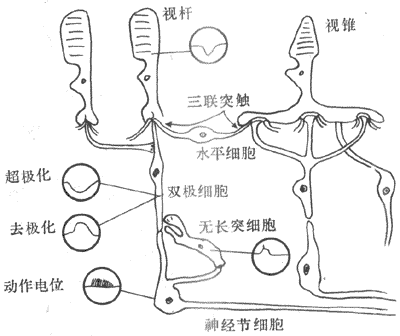
三、视网膜的结构和两种感光换能系统
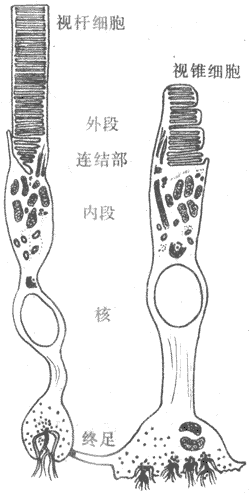
,,。,,;“”,,。,,、。,,。
(一)视网膜的结构特点
<>.-.,。,,。,,-。,;,,;,。,,。,,,,;,;。。,,,。,、、(-);,。,,。,。,。,,,;,,;。,,,。,。,,。,,,,,,。 <> ,,,,。,,,,。。,,。。 <>
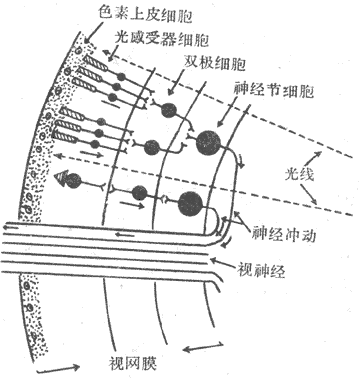 <>-
<>
<>-
<> <>-
<>-
(二)视网膜的两种感光换能系统
<>,。,,,;,,;,,,,,,()。 <>-:①,,;,;,;,,;,,。②,(),,,,“”,;,,;,;,。。③,,。,。④,(),,。
四、视杆细胞的感光换能机制
,,;,,(-)。,,。
(一)视紫红质的光化学反应及其代谢
<>-,,()()。,、α-,。,,。,()()(-),。 <>
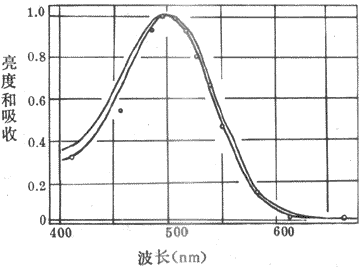 <>-
<>
<>
<>,。,,-(),()。,,,。,,,。,。
<>,,,。,-,。,,-,,-,;,。,,,;,,,;,,,,,;,,,,。,,()。,,。
<>-
<>
<>
<>,。,,-(),()。,,,。,,,。,。
<>,,,。,-,。,,-,,-,;,。,,,;,,,;,,,,,;,,,,。,,()。,,。
(二)视杆细胞外段的超威结构和感受器电位的产生
<>-。,-。,,,。,,,,。,;。。 <>
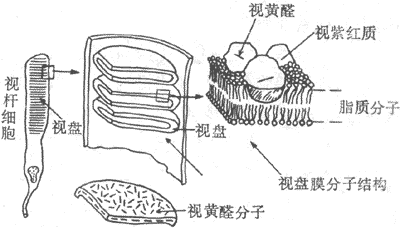 <>-
<>,,,-?/&;-,。,,[]+[/][]+[/],[]+[/][]+[/],[]+[/]。,,,,,,,(),,,。
<>,,(),-,,,,,[]+[/],,[]+[/],。,,;,。,[]+[/],。
<>,,()。
<>-
<>,,,-?/&;-,。,,[]+[/][]+[/],[]+[/][]+[/],[]+[/]。,,,,,,,(),,,。
<>,,(),-,,,,,[]+[/],,[]+[/],。,,;,。,[]+[/],。
<>,,()。
五、视锥系统的换能和颜色视觉
<>,,。,,。-,。,,。,,-。,。 <>,。-,,。、、、、、、,;,,-,。,,。,,。,,;、、(),。;、,。,()(),、、,,,。,;。,。 <>,,,,,,(,),,,,,,、、,。,,,(-),。 <>
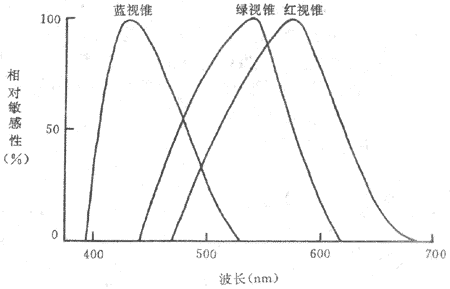 <>-
<>,。,;,,,。,;。,、。,,,,,。,。
<>,,,。,,(,,)。,。,。:。,,(,)。,。,,。,、,,,,,;。。,,,;(),,。,,。
<>-
<>,。,;,,,。,;。,、。,,,,,。,。
<>,,,。,,(,,)。,。,。:。,,(,)。,。,,。,、,,,,,;。。,,,;(),,。,,。
六、视网膜的信息处理
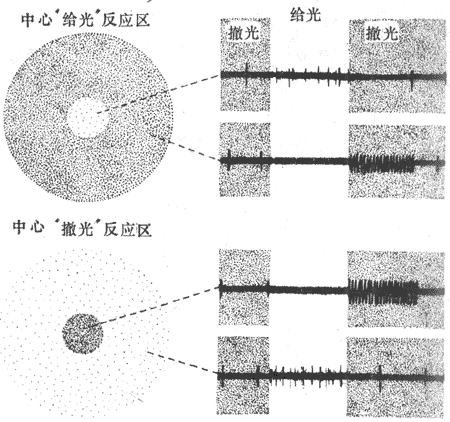
<>,,。,(,,),,;,,。,、,();,,,( ,)。,(),,();,,,,“”,(-)。 <>
 <>-
<>
<>,“”,()(),。,(),%。,“”();,。
<>,,-、--。--“-”;,;;,,(-,),-,,,,(-,)。
<>
<>-
<>
<>,“”,()(),。,(),%。,“”();,。
<>,,-、--。--“-”;,;;,,(-,),-,,,,(-,)。
<> <>-
<>--。,-,;-,,。,-,;-,。-,,,,,。,--,。
<>,,-,、();,。,,,“”,。“”,“”,,,、。
<>-
<>--。,-,;-,,。,-,;-,。-,,,,,。,--,。
<>,,-,、();,。,,,“”,。“”,“”,,,、。
七、与视觉有关的其它现象
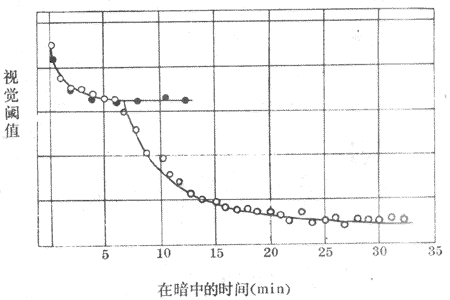
(一)暗适应和明适应
<>,,,,,。,,,,,。 <>。,,,、。-,,,;-,,。,。,;,。 <>
 <>-
<>。
<>·(,
<>,)
<>,。,;,。
<>-
<>。
<>·(,
<>,)
<>,。,;,。
(二)视野
,。(,,)。,,,,,。,。,,,,。,,,,。、,,。
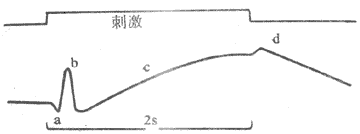
(三)视网膜电图
<>,,,,()。;。、(-)。,;,;,。,,。,,,。 <>
 <>-
<>-
(四)双眼视觉和立体视觉
,,“”。:,。,;,,,“”;,,,,,。,,;。