第一节 概述
<>-、。,。 <>。。,,,;。。,、,(-);。,,。,,,。、。,。 <>- <><><><><>(/)<><><><><><><>.-.<>.-.<><><><><><><><>α-<><><><><>.-.<>.-.<>、<><><><><><><>()<><>、、<><><><><><><><><><>.-.<>.-.<>[][/]<><><><><><><>()<><>、<><><><><>()<><><><><><><>()<><><><><><><><><><><><><><><><><><><><><>α-<><>、<><><><><><><>、、<><><><><><><>、<><><><><><><>、<><><>.-.<>.-.<><><><><><><><><><><><><><><><><><><><>.-.<>.<><><><><><><><><><><><><>.<>.<><><><><><><><>[][/]<><>
一、消化道平滑肌的特性
,、、,。,,;,。
(一)消化道平滑肌的一般特性
<>,、、,。 <>.。、,。 <>.,,,,。 <>.,。,、,;;。 <>.。,。。 <>.,、,。,、。
(二)消化道平滑肌的电生理特性
<>,,、。 <>.,,-—-,[]+[/],[]+[/]、[]-[/]、[]+[/]。 <>.;,,( ,)。,,/,/,-/。-,。 <>,,。。,,,;,,。,,。 <>,,。,,,。,。,,。 <>.:①,;②,[]+[/],[]+[/];③,[]+[/],,[]+[/][]+[/],,,。 <>[][/],,,,。 <>、:,。,,,,、(-)。 <>
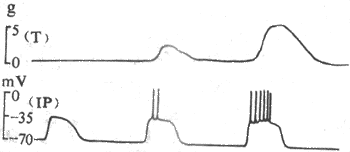 <>-
<>,
<>,;
<>,,
<>,: :
<>-
<>,
<>,;
<>,,
<>,: :
二、消化腺的分泌功能
<>-(-)。、。:①,,;②,;③,;④、,,。 <>,、,。-,,,,(-)。 <>
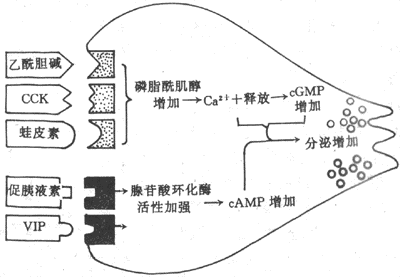 <>-
<>-
三、胃肠的神经支配及其作用
<>,(-)。 <>
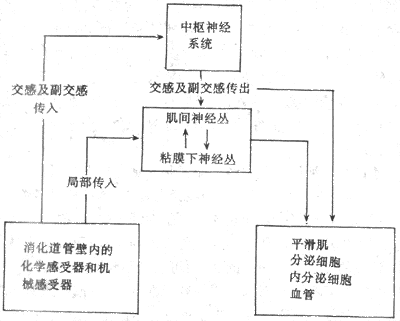 <>-
<>。();()。,,[][/],、。(),、、。,、,。
<>,。,、,,,:①;②;③。,,。,,。
<>
<>-
<>。();()。,,[][/],、。(),、、。,、,。
<>,。,、,,,:①;②;③。,,。,,。
<>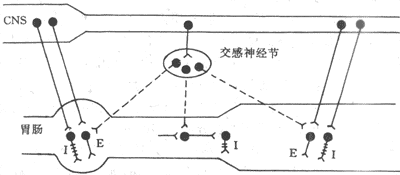 <>- 、
<>: : :
<>——: ----: +++
<>。,。,;,,,,(-)。,,()、、。,、,,、、(-)。
<>
<>- 、
<>: : :
<>——: ----: +++
<>。,。,;,,,,(-)。,,()、、。,、,,、、(-)。
<>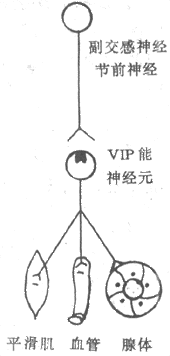 <>- ()
<>- ()
四、胃肠激素
, ,,( )。,。
(一)胃肠内分泌细胞的形态及分布
<>,,,。,,。,,(-)。 <>,,。、。,,,(-),。,,,。,,。 <>- 、 <><><><><><><><><><><><><><><><><> <>、、、<><><><>、<><><><><><><><><><><><><><><><><><><><>、、、、<><><><> <>
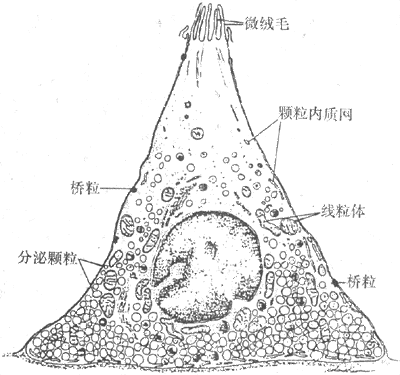 <>- ()
<>,。( -)。,、、、。
<>- ()
<>,。( -)。,、、、。
(二)胃肠激素的作用
<>,、。,。。 <>. 、、、、、-、。-。 <>- <><><><><><>[][/]<><><><>-<><><><><><>╂╂<>╂<>╂╂<>╂<>╂<>╂<>╂<>╂<>╂<><><>—<>╂╂<>╂<>╂<>╂<>—<>—<>—<>╂<><><>╂<>╂<>╂╂<>╂<>╂<>—<>╂<>╂<>╂╂ <>╂: ╂╂: —: <>.,,( ,)。,,。,,,,;,,。 <>:、、,、、、。 <>. ,( )。,、。。(,—),。,,,;,,,、。,,,、,。 <>,,。,,()。,()。-。 <>
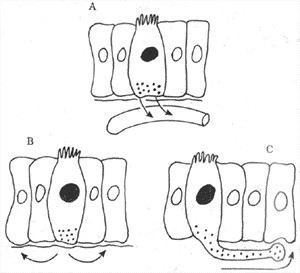 <>-
<>: :
<>-
<>: :
(三)脑-肠肽的概念
,,,;,。-( )。-、、、、。,,。
上一篇:第四节 呼吸运动的调节
下一篇:第二节 口腔内消化