第二节 白细胞分化抗原的应用
<>。,:(),;();();();()。,:();()、;()、;()。-。、。、、、。 <>- <><><><><><>< =><> 、、<> -/α()、/β/()<> ()、()、(/)、(/)、() γ()、γⅡ()、γⅢ()、εⅡ()α()< /><> -α()、-()、-()、()、 -γ()、-α()、-()、 -β()、<>-()、-()、-()、-()、<>()<><>、<><>-:<>-(/)--()、-()<>-(/)--()<>-(/)--()<>-()--()、-()<>--()、-()<><>-:<>-()-<>--()<>-(/)--()、-()<>-(/)--()<>-<>-(/β)-、-()<><>-:<>-(/)-、<>-(/)-、<>-(/)-、、<>-(/)-<>-(/)-<>-(/)-<>αβ(-/)-<>-β(/)-、<>-(/)-<>,(/)-<>ⅡⅢ(/)-、、、<>(/)-、、、、、<>αβ(/)-<>-β(/-)-<>/ β-<>/β-<>-α/(/)-<><>:<>-<>--()<>-Ⅱ<>-<> -Ⅰ<>-(/)--()、-()<>-/()<>-<><><>:()、()、、、、、 、、-(/)、<>:([][/]→[][/],)、()、( )、([]+[/],[][/]→[][/])、()、( )、、([][/]→[][/])、()<>:(?)、()<>:()、、<>:(,)<><><>()、()、(Ⅳ)、<>()、 (`)、()<><><>()、()、()、()
一、与T细胞识别、粘附、活化有关的CD分子
<>,,。,、,、()。( ,)(/)。,( )、, Ⅰ、Ⅱ,-(/)、、、/(-、-、-)、、、、-(),-()-()。 <>Ⅰ、Ⅱ“”。-、-、-、-、-、“”。“”。 <>
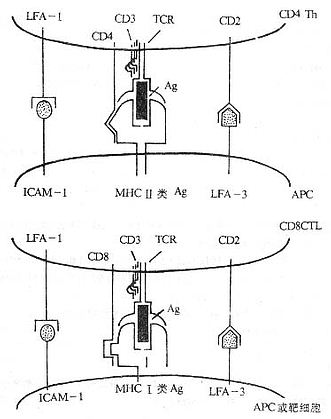 <>- ()
<>:
<>()(),---。
<>(),。
<>()。
<>(),、。
<>()、、、、-;-、-、-- 。
<>()。
<>(),,。
<>- ()
<>:
<>()(),---。
<>(),。
<>()。
<>(),、。
<>()、、、、-;-、-、-- 。
<>()。
<>(),,。
(一)T细胞受体
<>( ,) Ⅰ(Ⅱ),。,/,。 <>. 。,α、β、γδ。(),。~。,αβγδ。 <>()αβ:αβⅡ()Ⅱ αβⅠ Ⅰα~,β~。αβ()(),,。α、β,,~,,。α、β,。α、β()。~,α,β,γ、δε/,/,。α、β~(-)。 <>
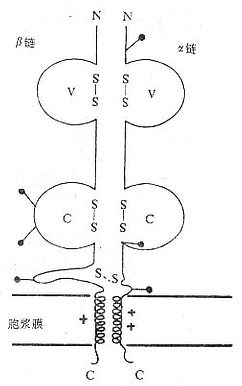 <>- αβ
<>() γδ:γδ、、,。γδ,δ,γ、δε。,γβ,δα。γδ,γδ。γ~,δ~,γ、δ。
<>α、β、γ、δ“”
<>. αβγδ,、、(-)。
<>- αβγδ
<><><>< =><>αβ<>γδ<>< =''> <>< => <><>~%<>.~%<>< =''> <><><> <>()、()<>< =''> <>< => <>+-<>~%<><%<>< =''> <><>-+<>%<>~%<>< =''> <><>--<><%<>~%<>< =''> <><><>%<>%<>< =''> <><><>>%<>-+()<>< =''> <>< => <><>(αβ)<>< =''> <>< = => < =>() <>(),Ⅱ;γδ。<>< =''> <><>()Ⅰ(、)<>< =>()<> <><>()(-、-、-、-、-γ)<>< =''> <>< =>()<>()γδ<>< =''> <><>()αβ。<>< =>() <> <><>()(、)<>< =''>
<>: , <>
<>,-+、+-、++--%、%、%%,αβ,--γδ。γδ:()、、-、-、γδ;()、γδ;()、()γδ;()γδ;()γδγδ,γδ;()γδ,γδ,γδ;()、γδ。
<>[](,),。,β,Ⅱ。:β+Ⅱ。、、,( ,),β.β,-βα。
<>+-,、,,、。
<>- αβ
<>() γδ:γδ、、,。γδ,δ,γ、δε。,γβ,δα。γδ,γδ。γ~,δ~,γ、δ。
<>α、β、γ、δ“”
<>. αβγδ,、、(-)。
<>- αβγδ
<><><>< =><>αβ<>γδ<>< =''> <>< => <><>~%<>.~%<>< =''> <><><> <>()、()<>< =''> <>< => <>+-<>~%<><%<>< =''> <><>-+<>%<>~%<>< =''> <><>--<><%<>~%<>< =''> <><><>%<>%<>< =''> <><><>>%<>-+()<>< =''> <>< => <><>(αβ)<>< =''> <>< = => < =>() <>(),Ⅱ;γδ。<>< =''> <><>()Ⅰ(、)<>< =>()<> <><>()(-、-、-、-、-γ)<>< =''> <>< =>()<>()γδ<>< =''> <><>()αβ。<>< =>() <> <><>()(、)<>< =''>
<>: , <>
<>,-+、+-、++--%、%、%%,αβ,--γδ。γδ:()、、-、-、γδ;()、γδ;()、()γδ;()γδ;()γδγδ,γδ;()γδ,γδ,γδ;()、γδ。
<>[](,),。,β,Ⅱ。:β+Ⅱ。、、,( ,),β.β,-βα。
<>+-,、,,、。
(二)CD3(T3、Leu4)
<>,γ、δ、ε、ζ、η,(-)。(-),。,。 <>
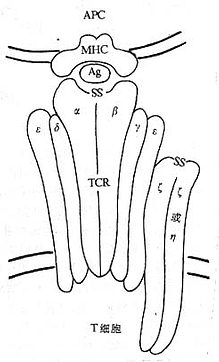 <>- /
<>,γ、δεα、β。γ、δε,,αβ,。ζ-ζαβ/γδεεαβ/γδεεζζ(αβ/γδεεζη)。ω( ,),/,。ω。
<>
<>- /
<>,γ、δεα、β。γ、δε,,αβ,。ζ-ζαβ/γδεεαβ/γδεεζζ(αβ/γδεεζη)。ω( ,),/,。ω。
<>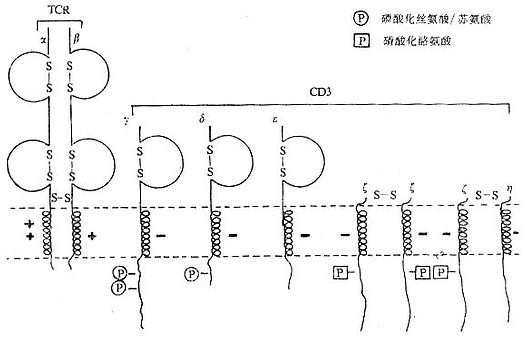 <>- /
<>.γ、δε γ、δε,,,。γ、δε(),,,。γ、δε/,α、δ、β、γ。γ、δε,。γ~,,,,,,。δ,,。δ,。ε,,。ε。
<>.ζη ζ()η(),γ、δε。ζη,,。,,,ζζζη。ζη。ζη,。,ζγ。,ζεⅠγ。
<>γ、δε,ζζ-ζ~%,~%ζ-η。/αβ/γδεεζζ。
<>,-,-、-α、-β、-γ-,,、。
<> 。、Ⅲ,。
<>- /
<>.γ、δε γ、δε,,,。γ、δε(),,,。γ、δε/,α、δ、β、γ。γ、δε,。γ~,,,,,,。δ,,。δ,。ε,,。ε。
<>.ζη ζ()η(),γ、δε。ζη,,。,,,ζζζη。ζη。ζη,。,ζγ。,ζεⅠγ。
<>γ、δε,ζζ-ζ~%,~%ζ-η。/αβ/γδεεζζ。
<>,-,-、-α、-β、-γ-,,、。
<> 。、Ⅲ,。
(三)CD4
<>ⅡⅠ,,。Ⅱ,Ⅰ。、“”。 <>. ,,,,,,。,。 <>
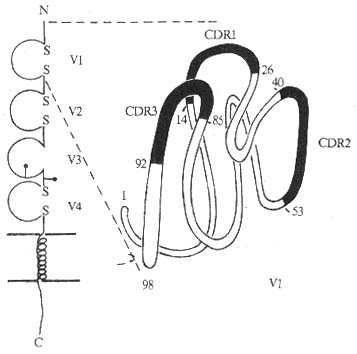 <>-
<>κ,(- , )。 ,,。Ⅱβ。,,,,,,( ),,,,。%,。,。。
<>. ,、、。,。。
<>. 、,/( / )。
<>():、ⅡⅡⅡ。。
<>():[][/],()。
<>,,。 :()、;()、;() ;()。+,:(),、,;()+ψⅡ,;(),,;()。,。
<>()( )、Ⅱ;()。,-、-()、-(),。(),,,(),、、、。()、Ⅱ。( )。
<>-
<>κ,(- , )。 ,,。Ⅱβ。,,,,,,( ),,,,。%,。,。。
<>. ,、、。,。。
<>. 、,/( / )。
<>():、ⅡⅡⅡ。。
<>():[][/],()。
<>,,。 :()、;()、;() ;()。+,:(),、,;()+ψⅡ,;(),,;()。,。
<>()( )、Ⅱ;()。,-、-()、-(),。(),,,(),、、、。()、Ⅱ。( )。
(四)CD8
<>. α、β,α,-;β,-。、、。αβ。α(α/α),γδ。。αβ,κ、λ~%,~%, αβ%。α、β,κ,,。- -,、、,-.、-.-.-.。 <>
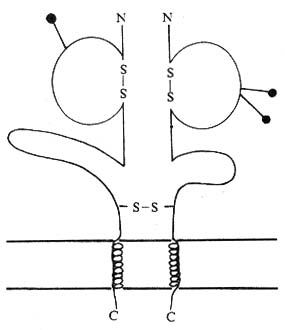 <>-
<>. 。γδα/α。α/α。
<>,/,()。、、、、、、,、、。
<>.
<>()Ⅰ。ⅠⅠ()Ⅰ。/( / ,/)。、 α,。
<>(),[][/]。。
<>,-Ⅱ。
<>-
<>. 。γδα/α。α/α。
<>,/,()。、、、、、、,、、。
<>.
<>()Ⅰ。ⅠⅠ()Ⅰ。/( / ,/)。、 α,。
<>(),[][/]。。
<>,-Ⅱ。
(五)CD2
<>. 、()、(-),,,,。,,(),,,,。%。%、~%(/)。 <>. <>():-,。-。。()、、;()-;()。,-3,--。-,--,-。-、、。,,。 <>()( ): :()[][/]-(),,[][/],[][/];()[][/],[][/][][/];()[][/](),[][/],[][/],[][/],。[][/][][/],Ⅱ-,-,-,,。-,。,,、。 <>,-,,,。
(六)CD58(LFA-3)
<>.- -( -,-)(),~,,,。,,。( )。 <>.- 、、、、、、、。 <>.- -,-(),;()、。,-,-。( ,)-、γⅢ。 <>-:(),、、;()“”,-“”。 <>[“”](,),,,。,(),()()。
(七)CD28
<>. .。,。,,,。%。-,。 <>. ,+~%,%+%+。+,++,+-。,。 <>
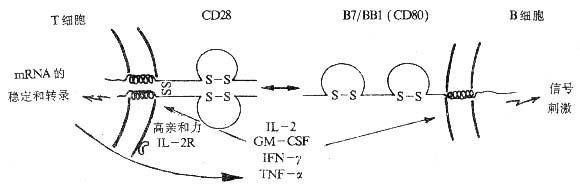 <>-
<>.
<>():、、、、,-、-、-α、-β、-γ、-,。,-。
<>()/(),-,。
<>-
<>.
<>():、、、、,-、-、-α、-β、-γ、-,。,-。
<>()/(),-,。
二、与B细胞识别、粘附、活化有关的CD分子
(一)BCR复合物
( ,),,,αβ,。,αβ,,αβ-。αβ。αβ,,(),。αβ,(-)。
(二)CD19
、,,。:();();()。,/α、β,,,/,,“”。
(三)CD21
<>( ,),。 <>[]( ,)、、。:()~( ,);();(),----。 <>. ,,,,,。,~,,-、-,,-,()-。。 <>. 、、,,、。 <>. <>():,(-),μ,;、-、;-,;;。,。。,,,。 <>():(),,[]+[/],。(-),()、,,,,,。 <>():,,。 <>():,Ⅰ。
(四)CD80(B7/BB1)
,,,,。,。,,-γ。,,、。
三、免疫球蛋白Fc段受体
、、、,。 , ,。γ、α、ε。
(一)FcγR(CD64、CD32、CD16)
<>.γ γγⅠ、γⅡγⅢ,。 <>()γⅠ():,,,~。、.、.γⅠ,[]-[/]~[]-[/],、。,。γⅠ、、,。γⅠ:~/,&;/,&;/。-γ、γⅠ~,-。 <>()γⅡ():,,,~。、·、。γⅡ,、,×[]-[/]。γⅡ,:~/。,γⅡ,γⅡ。 <>()γⅢ():~,,,~。、、、.。γⅢ、,。γⅢγⅢγⅢ:①γⅢ,,、,γⅢ,。γⅢζεⅠγ,γⅢγ,/γⅢζ。-βγⅢ。②γⅢ,“”,~γⅢ,γⅢ,。 <>
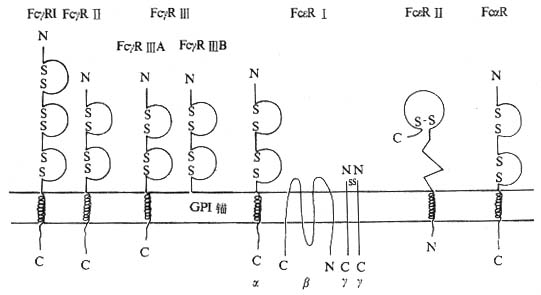 <>- γ、αε
<>.γ γ。
<>()-:γⅠ、ⅡⅢ,[]+[/],-。-γγⅠ。-γⅠ、Ⅱ、Ⅲ。
<>():γ,-γγⅠγⅡ,γⅠ,-γ,γⅡ,。-γⅡ。γⅢ。γⅠ、Ⅱ。
<>():,-、-,。-γⅡ。
<>():γⅢ,--γ,γⅢ。
<>- γ、αε
<>.γ γ。
<>()-:γⅠ、ⅡⅢ,[]+[/],-。-γγⅠ。-γⅠ、Ⅱ、Ⅲ。
<>():γ,-γγⅠγⅡ,γⅠ,-γ,γⅡ,。-γⅡ。γⅢ。γⅠ、Ⅱ。
<>():,-、-,。-γⅡ。
<>():γⅢ,--γ,γⅢ。
(二)FcαR(CD89)
α(),,,,,,,、、,、。α,×[]-[/]。。
(三)FcεR(FcεRⅠ、FcεRⅡ)
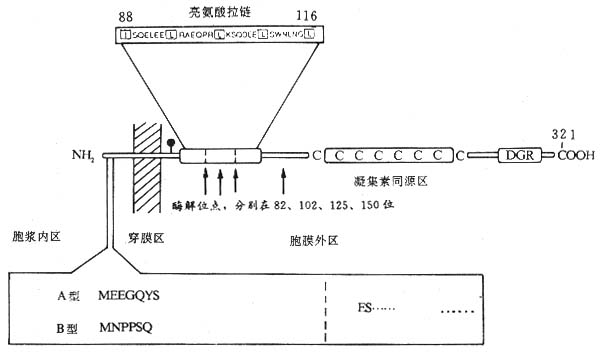
<>.ε εεⅠεⅡ,、。 <>()εⅠ:,[]-[/]~[]-[/],α、β、γ-γ。α,,,,γⅡγⅢ,ε/ε;γⅢ;。β,,,,βαγ-γ。γ,,,,γ,γεⅡ。γⅢ()ζεⅠγ,εⅠγζ。εⅠ。 <>()εⅡ():,,,Ⅱ,。,,~,~,,,、、、,,。,。、~(- -)。εⅡ()-。εⅡεⅡ,(-)。εⅡ,;εⅡ、、、、、、、、、,,εεⅡ。-、、εⅡ,,。-γ、-β、、。 <>.ε <>()εⅠ:εⅠ,,、/εⅠ,[]+[/],,、、,Ⅰ。 <>
 <>-
<>()εⅡ:εⅡ,,-()。( . , ),(-)。(),。,,,。,,-,,εⅡ。,,,εⅡ-~- ,εⅡ,,,。(-),。
<>、,,。 ,。(-),,。
<>()
<>-
<>()εⅡ:εⅡ,,-()。( . , ),(-)。(),。,,,。,,-,,εⅡ。,,,εⅡ-~- ,εⅡ,,,。(-),。
<>、,,。 ,。(-),,。
<>()
上一篇:第一节 白细胞分化抗原的分类
下一篇:参考文献