第一节 补体固有成分的分子结构及功能
,(-,、、)。,,、、,。
一、C1分子
。+。。,。
(一)C1q
<>()γ。,、、。、、,。、、,、,-,。,(--,)。。、。,。。,。/,。,(-) <>
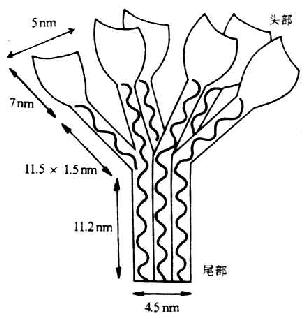 <>- ()
<>。,。(-),,。,,。:&;&;&;。
<>、,、。,。、、.-.。
<>- ()
<>。,。(-),,。,,。:&;&;&;。
<>、,、。,。、、.-.。
(二)Clr和Cls
<>,()。 。,。,(),,()。 <>,,。 <>
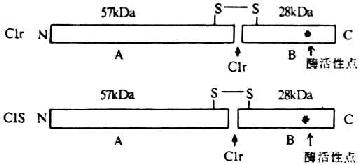 <>- /
<>+“”,(-)。(-)。。,,、,,。,(-)。。,。,,,,,。(),()。
<>
<>- /
<>+“”,(-)。(-)。。,,、,,。,(-)。。,。,,,,,。(),()。
<>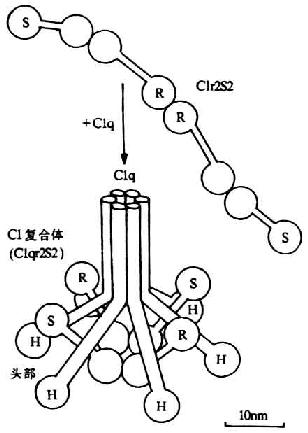 <>- (、)()
<>,。-,。
<>- (、)()
<>,。-,。
二、C4分子
<>,,α()、β()γ()(-),α。α。α-(-),。(.),,,,,%。α`,,。,[][/],(-)。 <>,/,。,()(),、,。,。 <>--。[]*[/][]*[/],,()[]*[/][]*[/]。、、Ⅲ。 <>
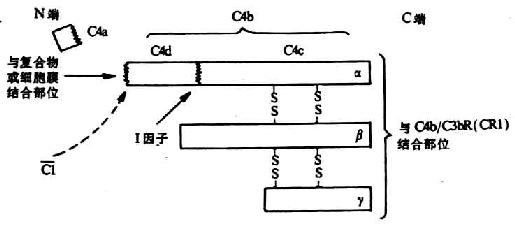 <>- ()
<>- ()
三、C2分子
<>,。,,(-)。,(-),,。,,,。,,()()。,,。 <>
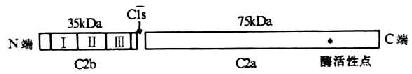 <>- ()
<>:Ⅰ、Ⅱ、Ⅲ
<>,。,;,( ,)。()。
<>- ()
<>:Ⅰ、Ⅱ、Ⅲ
<>,。,;,( ,)。()。
四、C3分子
<>,,。α、β,,,α,β(-)。,.-./。,α(---)。,,,“”。α-(-),(),。,α`,(-),。,(--)。,,(-),。,,,,(-)。(、)()。 <>(),()( ,,(-)。,,。,。 <>。,,,,,。。,,、()。,:()[()()],();();();(),,,(),、,。。、,。 <>,。,。,,。 <>
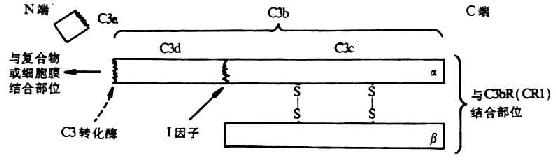 <>- ()
<>- ()
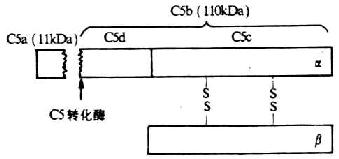
五、C5分子
<>()。α、β, ,α,β(-)。,。-。,α,,。,,。,。 <>,。:():,,。,,。():、,。, ,,。():,,,。,。():,,-、-、--α,。,。-。 <>
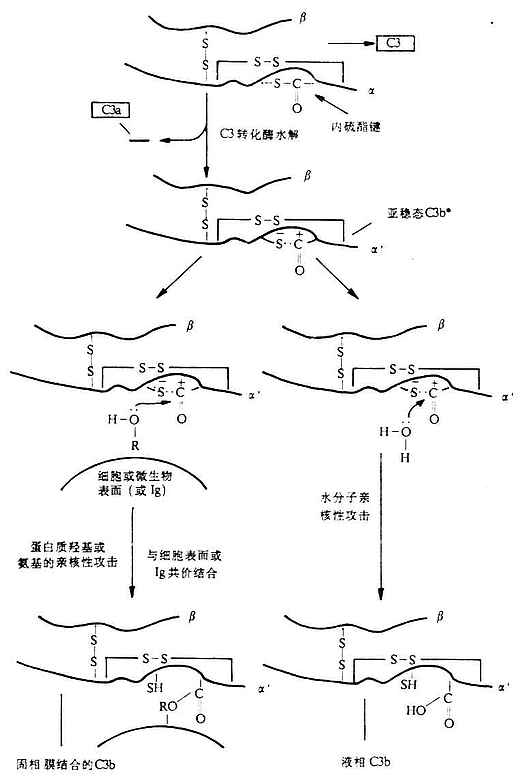 <>- ()
<>
<>- ()
<>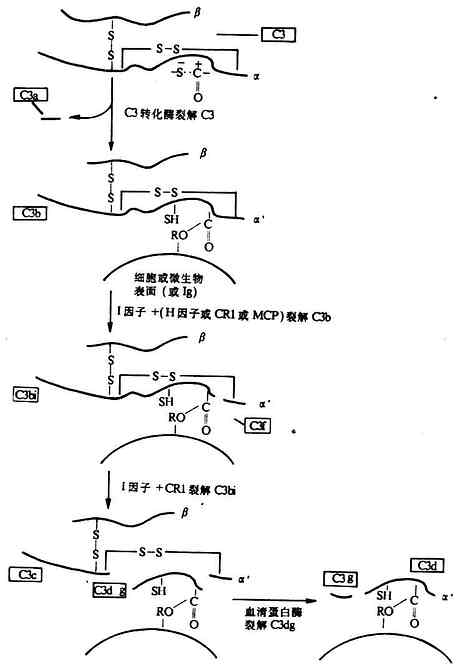 <>- ()
<>
<>- ()
<> <>- ()
<>- ()
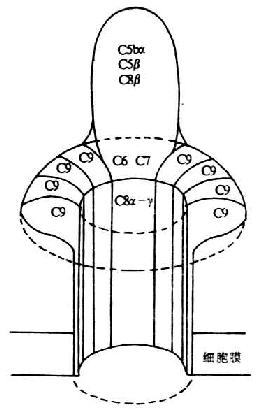
六、C6和C7
<>,,。。.%。,,,,-%。,。(),,。 <>()、、Ⅰ(-),。,Ⅰ()。,,。,。,-,。, <>,。。,,。,,。,(),。 <>。,(),。,。
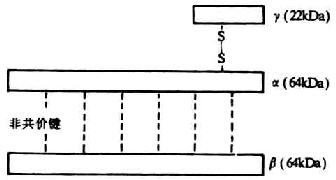
七、C8分子
<>α、β、γ,。αβ,γ。αγ,αβ(-)。-。αβ(-),,()。αβ。β,,。-β。 -,,α,.,,。-,。, -,-,-。 <>
 <>- ()
<>,αβ,γ。β,,,。αβ,%,%
<>- ()
<>,αβ,γ。β,,,。αβ,%,%
八、C9分子
(),,。,。,。 -。,,℃, &;-,。-,、(-)。,。,。αβ,-、。,。,。
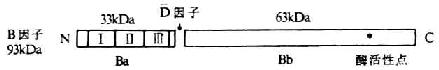
九、B因子
<>( ,),。(%),。。,。,(-)。[]+[/],,。()()。,, ,。 <>
 <>- ()()
<>
<>- ()()
<> <>- ()
<>Ⅰ、Ⅱ、Ⅲ
<>,。 (),(),。,。,,。
<>- ()
<>Ⅰ、Ⅱ、Ⅲ
<>,。 (),(),。,。,,。
十 、D因子
,,。(-μ/),。。()--,。,,(、、)。
十一、P因子
<>(),。,:(%)、(%)(%),:&;&;。(),,。 ,,。,,,。,。 ,。,( ,) , ,-。 <>-。 <>- <><><><><>(μ/)<>()<>()()<><><><><><><>:<> <>、 <><> <> <> <>:<> <> <><> <> <> <>:<> <> <><><><><><><>,<><><><><><><>,、<><><>-<><>α:<><><><> <> <> <>β:<> <> <><> <> <> <>γ:<><>、,、、、<><><><><><><><><> <> <> <> <><>,、<><><>-<><>α:<><>、<><> <> <> <>β:<><>、、,、<><><><><>α:<><>、、<><> <> <> <>β:<> <><><> <> <> <> <><><><><><><><> <>,<><><><><><> <><><><><><>α:<> <>,<><> <> <> <>β:<> <> <><> <> <> <>γ:<> <> <><><><><><> <><><><><><><><><><> <> <> <> <><>,、<><><>~<><><><>,<><><><><>,<> <>、 <>::;:;:
上一篇:第五章 补体的分子生物学
下一篇:第二节 补体调节分子的结构及功