第二节 T淋巴细胞及其亚群
、( ),。
一、T淋巴细胞在胸腺中的分化
(一)T细胞在胸腺分化过程中的表型改变
<>,,-,。 <>,、:()( ,),“”( );()(-、--)(、);()(-、-)。,、、。,。 <>. ,αβ,αβ。/,。 <>./ (,-),,。,αβ(γδ),/,,( )。 <>. 。-。 <>- <><><><>< => <>%<>< =>< =>+----<>< =>~<>< =>↓<> <>< =>++--+<><>< =>↓<> <> <>< =>++++++<><><>< =>↓<> <> <>< =><>+++<>++-<><><><>-+<>++<> <> <><>↓<>↓<> <> <>< => <> <>-,。 <>-,,,,-,。-。、、、-,。--。,-。-,。,-,-,-,。-,-,-。-。-,-,-,,:-.-.。-β。 <>(-,),-,。-、--。、、、,。/,。
(二) T细胞在胸腺中的选择
<>、。()。 <>. Ⅱ,,( )。 (),; Ⅱ,。+- ,+- Ⅱ,。 <>.( ) ,、。( ,) Ⅱ, Ⅱ。,( ),+--+,。 <>,:(),()( );();(),;()、()。
(三)T细胞在胸腺中获得MHC限制的能力
<>,,。(-[][/] -[][/])-[][/]-[][/]。(-[][/]-[][/])(-[][/])-[][/],-[][/],-[][/](-)。 <>- <><><>< =>< =>< =>< =>< =>+<><>-[][/][][/]<>-[][/]<><>-[][/]<>-[][/]<>-[][/]<><>++<>++<><>-[][/]<>-[][/]<>-[][/]<><>++<>-<><>-[][/]<>-[][/]<>-[][/]<><>-<>++
(四)成熟T细胞的膜表面分子
<>,,,,。()、()。 <>. “”,“”。 <>():。( ,),,,,( ),。:[][/]、[][/][][/]。[][/],[][/]。[][/]( -,-),,。[][/],。[][/]。 <>():,。γ、δ、εη,/。,。 <>():( / ),。,(),。,,,( ,)。Ⅱ,Ⅱ。 <>- <><><><><><><>< =><><>()<>()<><><><><><><><><><><><><><>,<><><>////<><><><><><><><><>/<><><>/<><><>/,<><><><><><>,<><><><><> <>-α,,<><><><> <><><><><><> <> <>++, <>():α、β,、α。( ,)( ,),。 。,() 。 <>.() Ⅱ, , Ⅱ,。 <>.( ) ,。 <>()( ,) 。αβ,αβ,γδ。,αβ()()。αβ-----,,,。 <>():(),。 <>():,,。,( ,)( ,),。 <>- 、 <><><><> <><><><>%<>~<>~<><><> <> <><> <>++<>++<><>Ⅱ<><>++<><>、、<>-<>++<><><><>-<><><> <> <><><>-<>++<><><>++<>-<><><>++<>-<><>()<>++<>-<><><>++<>-<><><>++<>-<><><>++<>++<><><>-<>++()<><><>-<>++<><>()<>++(+)<>-<><>(,)<>-<>++ <>():()。,。。(,),(,)。,,( ),。,,,,。 <>(),,。、,。 <>()( ,):,,-(-)、-、-、-、-、-、-、-、-、-α、--β。,-β,-α,β、γ-。 <>- <><><><> <> <>()<><><><><> <><>α---<>,<><>()<>()<> <>α--<><><><> <><>---<><><>()<>( )<> <><> <><><><><>β----<>、<><>()<>()<> <><><><><> <><>--<><><>()<>()<> <>(β-)--<>(+)<><> <> <> <>--<>,<><><> <><>----<>,<><>()<>()<> <> -<> <>、,、、、、,、,、、,-、。、。
二、T淋巴细胞亚群
+--+。。
(一)CD4阳性细胞群
<>. ,,,-。 <>-、-γ、,、-、-α-,-、-、-、--;-α、-、-、-、-、-、-(,)-,-、-γ。。,。,-,-。-γ-。-,-γ。,--,-(),,,。,。--,,-,,。-γ,。,-、-γ,。()Ⅱ,-γφ,-φ。。,++,-γ,--γ++。,-%,%+-γ-。 <>- <><><><> <> <><><>< =><>-<>+<>-<><>-γ<>+<>-<><><>+<>-<><>-<>+<>+<><>-α<>+<>+<><>-<>+<>+<><>-<>-<>+<><>-<>-<>+<><>-<>-<>+<><>-<>-<>+<><>-<>-<>+<><><> <>+<>++<><><> <>-<>+<><><> <>-<>+<><><> <><><><><> <>φ<>,φ <>( ,)[][/],++-,。[][/],,(Ⅳ)。 <>- [][/] <><><><><> <><><><>()<>、、,<><> <>()<><><> <>()<> <><> <>-γ<> <><><>()<>、<><><>-<>、<><> <>-γ<><><><>()<><><><>()<> <>: <>: <>: <>: <>: <>: <>. 、、。-。 <>():、。-(,Ⅱ),,。,,,();-,。+,,;+-。,。 <>,/(-),-,(β)+。+。 -。。++。 <>():。、。;、、。,,、Ⅱ、、,,、。,。 <>- <><><><> <><><><> <>+<>+<><><><><><><>+<>-<><><><><><>(、)<>+<>+++<><><>+<>+++<><><>+<>+++<><> <>+++<>+<><><>-<>+++<><><>+++<>+++<><><>+++<>+<><><> <> <><><>±<>+++<><><>+++<>-<><>()<>±<>+++<><><>()<>() <>():,( ,)。。Ⅱ,Ⅱ。,。
(二)CD8阳性细胞群
<>+(++)(+-)。-%,。 <>.() +-++。-+、-+、-+/-+。 <>():()(-),(), ,/,-,-,-、-、-,-、-、-、-γ,,( )。,() 。 ,“”。( , )。-,,。 <>():,:①-/-、-、-,-(-);②/-(),;③/。 <>():。 <>①():(- ,)、( )(),(- ),,-,.,--%,。,,,[]+[/],,(),-,。,。[]+[/]、,+(),,。,。( )、( ,),。 <>
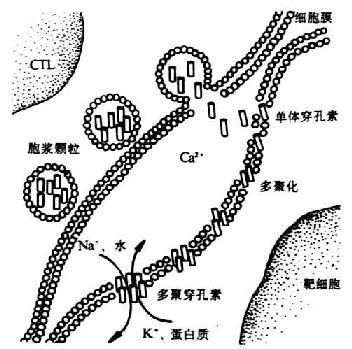 <>-
<>②( ):,-( )、-( ),,。
<>③(,):-β(-β),,,“”。
<>. ( ,),,、。
<>():( , ),。,,。,,。-,。。
<>-
<><><><>()<>---→+++<>< =><>< =>()---------------------------------→+<>< =><>< =>()-→-→-→+<><> <> <>< =>()-→-+ ----→-→+++<>< =>
<>,+-+(-)。+-+-。,、。
<>():、,、。,。(,)(,)→(,),( )→(,),(),,,。,、、,(- ,)。,,,。
<>-
<>②( ):,-( )、-( ),,。
<>③(,):-β(-β),,,“”。
<>. ( ,),,、。
<>():( , ),。,,。,,。-,。。
<>-
<><><><>()<>---→+++<>< =><>< =>()---------------------------------→+<>< =><>< =>()-→-→-→+<><> <> <>< =>()-→-+ ----→-→+++<>< =>
<>,+-+(-)。+-+-。,、。
<>():、,、。,。(,)(,)→(,),( )→(,),(),,,。,、、,(- ,)。,,,。
上一篇:第一节 B淋巴细胞的分化和表面标志
下一篇:第三节 自然杀伤细胞