第二节 抗原的处理与递呈
一、抗原的捕获与处理
辅佐细胞可通过多种方捕获抗原,例如吞噬作用(对同种细胞或细菌等大型颗粒)和胞饮作用(对病毒等微小颗粒或大分子)等。这种吞噬和吞饮作用无抗原特异性,可能的识别机制在于吞噬细胞与被吞噬颗粒之间的表面亲水性差异。另外还有受体介导的内摄作用,这是弱吞噬力的辅佐细胞捕获抗原的主要方式,例如B细胞可借助抗原受体(表面免疫球蛋白)与相应的抗原特异性结合,并将抗原内化处理。这些捕获方式与中性粒细胞的吞噬作用(详见第八章)相似。
抗原处理(antigenprocessing)是指辅佐细胞将天然抗原转变成可被T[XB]H[/XB]细胞识别形式的过程;这一过程包括抗原变性、降解和修饰等。例如细菌在吞噬体内被溶菌酶消化降解,将有效的抗原肽段加以整理修饰,并将其与MHCⅡ类分子相连接,然后转运到细胞膜上。
可与MHCⅡ类分子结合的都是蛋白性抗原;多糖和脂类不易于MHCⅡ类分子连接,难以被T[XB]H[/XB]细胞识别,因而多不是良好的免疫原;但有时可以诱导抗体性免疫应答。
二、抗原的递呈
抗原递呈(antigenpresentation)是辅佐细胞向辅助性T细胞展示抗原和MHCⅡ类分子的复合物,并使之与TCR结合的过程。这个过程是几乎所有淋巴细胞活化的必需步骤。
抗原递呈之前,经处理后的抗原肽段已经连接在MHC分子顶端的槽中,这个复合物便是TCR的配体。TCR与配体结合的精确模式尚未清楚,一个合理的说法是TCR中α和β链的V段接触MHC分子的α螺旋(形成MHC分子顶端槽的肽段),使高可变的连接部(V-J及V-D-J)与抗原肽段相结合。这样保证了TCR识别抗原的特异性。
超抗原(例如细菌毒素等,详见第二章)的递呈有独特的模式,它不需要胞内处理,可以直接与MHCⅡ类分子结合。超抗原不结合在MHCⅡ类分子的顶端槽中,而是结合在槽的外侧;与TCR结合时,不结合其α链,只结合β链的V节段。超抗原对TCR和MHCⅡ类分子的结合都非常牢固,象一支双向钩子将T细胞和辅佐细胞紧紧地连在一起,很容易使T细胞活化。另外,任何超抗原都只与含特殊β链V节段的TCR结合,这样的TCR约占外周T细胞总数的1%~10%,这一数字远远大于任何普通抗原所能识别的细胞数;所以某些产毒细胞感染时,容易发生急性期素休克综合征,就是超抗原刺激的结果(图7-1)。
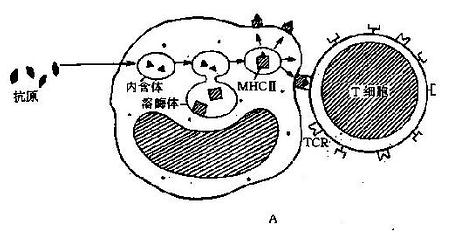
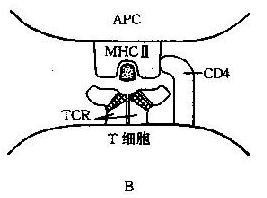

图7-1抗原递呈示意图
A:APC的抗原处理和递呈;B:抗原、Ⅱ类分子、TCR与CD4的相互关系
C:超抗原作用示意图
需要强调的是,TCR只识别自身或与自身相同的MHC分子,这就是MHC对TCR识别的限制性。这种限制性是T细胞在胸腺内发育成熟的过程中,由阳性选择作用所规定的。
除了辅佐细胞外,体内表达MHCⅠ类分子的细胞都可向Tc细胞递呈抗原,自身成靶细胞而被Tc杀灭。这类递呈作用是重要的免疫效应方式之一。
三、辅佐分子
TCR虽能与抗原-MHC复合物特异性结合,但这种结合力很弱(超抗原除外),不足以将辅佐细胞和T细胞连接在一起,还必须有额外的辅助力量使两类细胞密切接触。这些力量来自辅佐受体(accessoryreceptor)和协同受体(coreceptor)。
1.辅佐受体T细胞表面有多种粘分子(adhesionmoleule),例如CD2和LFA-1等。T细胞与辅佐细胞最初接触的能力来自这些粘附分子与相应配体的相互吸附,例如T细胞CD2与辅佐细胞CD58(LFA-3)的吸附。当TCR与抗原-MHC复合物相互识别以后,会加固粘附分子之间的作用。另外,辅佐细胞上的FcR、CR和丝裂原受体等也可以起到辅佐受体的作用。
2.协同受体T细胞亚群的标志分子CD4和CD8邻近TCR,对MHC分子有亲和力,可以分别直接结合MHCⅡ类和Ⅰ类分子。CD4和CD8之所以称为协同受体,不仅可增加T细胞与APC的接触,且在TCR转导刺激信号方面也起到一定的辅助作用(图7-2)。
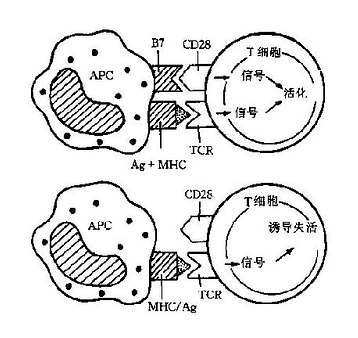
图7-2辅佐分子作用示意图