第二节 封闭群动物的特点及应用
一、定义和和概念
(一)定义
封闭群动物是指一个种群在五年以上不从外部引进新种,仅在固定场所的一定群体中保持繁殖的动物群。
(二)概念
封闭群(Closed colony)的正确概念是指引种于某亲本或同源亲本的动物,让其不以近交形式,也不与群外动物杂交而繁衍的动物群,目的是要求整个群体尽量防止近亲交配而保持着遗传变异,也就是说既保持群体的一般特性,又保持动物的杂合性。至于个体间差异的程度因引种来源的不同而有不同,如引种于一般杂种,则个体间差异就大;如果引种于有近交历史的动物,则个体间差异就小。
由于封闭群体本身的特点,加上客观上具有各种各样的相似存在形式,就使封闭群这个概念很混乱。有人叫“非近交群”(Non-inbred colony),有人叫“非近交系”或“非近交品系”(Nou-inbred),甚至有人叫“远交系”、“远交株”或“远交动物”(Outbred animals)。实际上这些叫法并不是指真正的“封闭群”,而只是通常供应使用的、保持于各种饲养系统的非近交群体而已。特别是“远交”一词很容易使人理解为遗传学上的“异系交配”。因为“远交”在遗传学上是指遗传上不相关的动植物的杂交,或没有相近亲缘关系的不同家系的两个个体的交配,甚至把不同特种、不同变种的交配都看作是异系交配。而实验动物的封闭群,其个体之间并不是没有亲缘关系,甚至是很亲近的或有一定的近交。所以封闭群不应叫成上述这些名称,它们之间更不能划等号。
ICLA在1963年和1964年曾规定:“不从外部进行引起的群体为封闭群”,这是最基本的。日本实验动物研究会于1973年又进一步规定:“五年以上不从外部引种,只在一定的群体中进行繁殖,为经常提供实验动物而进行生产的群体叫做封闭群”。
实际上对封闭群的全面理解应该包括封闭年限、群体大小及繁殖结构等。一般认为封闭群是指引种于某亲体或同源亲本的动物。
二、封闭群的分类
封闭群按其来源和遗传背景不同,可分为以下两大类:
1.来源于近交系的繁殖群及其子代,不用兄弟姐妹交配方式保种进行生产的实验动物。
2.来源于非近交系,不是以培育近交系为目的而生产的实验动物群。
如系来源于近交系的封闭群,虽然对繁殖群的的大小不作特殊规定,但应采取在群体内不产生隔离状态的方法繁殖。
如系来源于非近交系的封闭群,应经常保持“群体的有效大小”,一般为50只以上,同样应采取群内不产生隔离状态的方法繁殖。
上述两种封闭群,除了在选择时应考虑繁殖力外,均不采用特殊的淘汰方法进行选种。
三、封闭群动物的应用
目前国内外实验动物的使用,以小鼠为例,大部分是近交系和封闭群。而从使用量上来看封闭群远远超过近交系,这是因为近交品系繁多,又不易大量生产,往往仅适用设备条件较好的研究机构或专门科技人员亲自保种使用,大大限制了其使用范围。而封闭群因为有杂合子并避免了近交,故能保持相当程度的杂合性,从而避免了近交衰退的出现。所以,其生活力、生育力都比近交系强,具有繁殖率高等遗传学的特点,因此封闭群动物可以大量生产,作为鉴定实验用。例如ddN小鼠、NIH小鼠、LACA小鼠、Wistar大鼠以及目前各研究所长期自行繁殖的瑞士种小鼠、青紫兰兔、新西兰白兔、大耳白兔、豚鼠等均属此类。这类动物在生物制品和化学药品的鉴定上,其反应稳定性远远优于市售动物,特别是作为热源质试验的家兔更为明显。进行各种筛选性实验时,选用封闭群动物有一定优点,因为在这群动物中有的可能有近亲关系,有的可能没有,而保持一定的遗传差异。因而对各种刺激的反应有强一些的,也有低一些的,但其平均的反应性有一定稳定性,故要观察筛选某一药物的初步疗效时,封闭群动物就可以反映综合的平均疗效。
然而,对于封闭群的研究,无论在理论上还是实践上,无论在国内或国外,都落后于近交系。其原因在于封闭群是属于群体遗传学理论范畴,不仅群体遗传学本身产生较晚,而且该理论又不能机械地套用于封闭群。到目前为止,对封闭群动物所进行的研究工作几乎都只限于小鼠和大鼠,而且报告也甚少。
四、培育方法及注意事项
封闭群动物的培育方法较为简单,只要不引进新品种,让其自行繁殖就可以,当然要避免近亲繁殖。但是长期保持一个封闭群就必须要控制各种条件,采取一定的措施。保持封闭群的主要目的的是为了减少群体内的遗传变异,使整个群体的性状、特征能持续稳定不变,而且不产生性质不同(变异)的个体或小群体。培育时应注意下列规则:
(一)群体封闭后应维持五年以上
为什么把小鼠封闭群有效时间定为五年?其理由如下。假如将两个近交系进行交配,培育出一个新的群体(杂交群)。此时,把其中一个近交系的基因型假定为A/A,另一近交系为a/a,则两者杂交以后所产生的F1基因型必定是A/a,然后让F1进行随机交配,从理论上讲,F2代基因型分离的比例A/A为25%,A/a为50%,a/a为25%。再让F2进行随机交配,生下F3,其基因型的比例和F2一样,不发生变化。但实际情况不会如此准确,因为还有其它许多因素(如繁殖率的高低)的影响,假如具a/a基因型的个体比A/A及A/a基因型的个体繁殖率低1/2的话,此时这三种基因型出现的频率就各不相同(如图5-1所示),即随着代数的增加,a/a型频率逐代减少,而A/A型的频率则逐代增加,也就是说,在起初10代(从1~10代),各基因型频率,都有变化,达到15升后才趋于稳定(几种基因型频率平行发展)。这虽然是理论的推导,但已用于实践,故目前暂定15代,也就是说经过15代后,该群体的基因型频率才能达到相对稳定。以小鼠为例,一年大约繁殖3代,要达到15代,就需要5年时间。因此,5年确定为小鼠封闭群的最低年限,但这种年限只能用之于小鼠,如豚鼠和兔等世代间隔长,封闭群达到稳定的年数,就比小鼠的更长了。
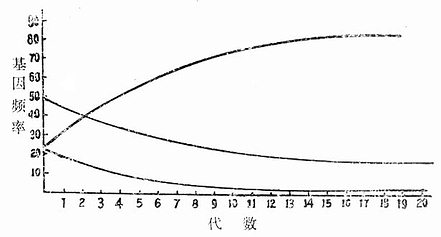
图5-1 不同繁殖率基因型个体的基因型频率变化:在A/A为35%,
A/a为5%,a/a为25%的群体中a/a的繁殖率为A/A和A/a的1/2时,各基因型频率的变化。
(二)防止产生群体内的隔离状态
封闭群体,不要在隔离状态下进行繁殖(交配)。上面所讲的是随机交配时的情况。如果交配不随机(如一封闭群隔离饲养,每处的雌雄只数都不多),而是兄弟姐妹之间交配,则该封闭群就会分化为许多近交系,从而使该群体的基因型结构发生变化,结果不但破坏了封闭群固有的遗传结构,而且将使封闭群分化成几个近交系,如近交系之间再进行杂交,虽可提高该群体的繁殖率,可是封闭群原有的特点将因此丢失。
假如将某封闭群分在两个饲养间饲养,各饲养间又独立地自行保种,这样就使群体有效大小减少一半,随之近交系数也将提高,从而在这两个饲养间产生基因结构(型)变化的可能性也就大了。因为,基因结构的变化原则上趋向于不随机交配的群体一方。如果经过若干代之后就会变成两个完全不同的封闭群。即使是各饲养间的群体量很大,但相同基因结构的微小变化是不可避免的,因此经过几十年代后,这些微小差异积累起来,也同样产生上述结果。有时虽然在同一饲养的保种,但采取的方法与上述两饲养间的作法相同,结果也会产生两个封闭群。
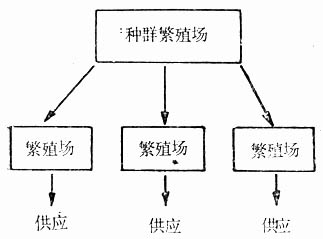
图5-2 繁殖场较多时,为避免各繁殖之间产生隔离生产的方法
总之,若同属于一个封闭群,则留种对必须在整个群体内普遍地选择,并进行随机交配。如果繁殖场有两个以上都生产同一品系的封闭群,为避免隔离状态下的配种,可采用图5-2方式繁殖生产。即由一个种群繁殖提供雌雄种鼠,再分配给各繁殖场,进行随机交配生产,然后供应使用。
(三)“群体有效大小”应保持在50只以上
当采用图5-2这种生产方式时,应特别注意种群繁殖场的“群体有效大小”,要达到足够大。如果封闭群过小,必然引起群体内近交系数上升,而致群体内各种基因的物质纯度提高,这是与封闭群的性质所不相容的,是封闭群最忌讳的。如群体只有雌雄两只组成,就只能进行兄妹交配,结果会育成近交系。如果群体有10只组成,这时虽然1、2代不进行兄妹交配,但在2~3代后,不可避免地要进行堂兄弟和堂姐妹之间的近亲交配,从而也能使群体的近交系数上升,结果使群体内基因结构改变。为了防止这种现象出现,必须规定群体的大小。
“群体有效大小”是群体遗传学的术语,有固定的含义。即在进行动物的繁殖生产时,为表示其群体规模大小,常以雌雄种的数目来表示。这个数目,通常的含义可定义为该群体的大小,但是当考虑每代之间的遗传特性时,这种表示方法就有问题了。比如,雌雄种的数目虽然相等,但在留种时如来源分布不均时,情况就大不一样(见表5-1)
表5-1 由10对公母鼠组成的不同群体的有效大小
| 双亲号/留种仔数/留种方式 | 12 3 4 5 6 7 8 9 10 | 计 | 群体的有效大小 |
| A | 2 2 2 2 2 2 2 2 2 2 | 20 | 40 |
| B | 1 3 0 3 0 3 4 1 2 3 | 20 | 20 |
| C | 0 4 0 3 4 0 0 0 5 4 | 20 | 12 |
| D | 0 12 0 0 0 8 0 0 0 0 | 20 | 4 |
表中所示为10对种动物,采取A、B、C、D四种不同的留种方法。从中不难看出,四种方式虽都是在10对亲代所生的仔代中选种,但是所选出留种仔数,在A组是来自10对种鼠,B组是来自8对种鼠,C组来自5对种鼠,而D组则只是来自2和6号两对种鼠。四组选出的留种仔数分别都是20只,群体的有效大小从外观上似乎相同,但其基因结构则完全不一样。为了比较这种差异,就采用了“群体有效大小”这一标准来进行衡量。
“群体的有效大小”是指一个自然繁殖的群体所包含的雌雄动物数目,是为下一代留种时,能从中随机选种,并随机交配以繁殖后代的数目,并以此作为衡量的尺度,以计数被测群体的有效大小。表5-2的B组所列即大致与所谓理想大小接近,故其有效大小定为20,A组则为40,正好为D组的10倍,C组为12,D组为4。如果采取A组的形式进行繁殖,必须非常仔细选种才能达到,因此非常麻烦。B组实际是采服随机选择的一例,因此操作较简便,只要稍微留心选种即可,但是如果不留心,很可能会出现C组或D组那种情况,即仅仅从几只母种所生的仔代中选择,势必造成群体有效大小的变小。
降低“群体的有效大小”的另一主要原因是种鼠的雌雄数不同。雌雄鼠的性比越大,其“群体的有效大小”就变小(可见表5-2)。
表5-2 群体大小相同、雌雄种的比不同时的“群体的有效大小”
| 雌 种 数 | 雄 种 数 | 群 体 大 小 | 群体的有效大小 |
| 10 | 10 | 20 | 20.0 |
| 12 | 8 | 20 | 19.2 |
| 14 | 6 | 20 | 16.2 |
| 16 | 4 | 20 | 12.8 |
| 18 | 2 | 20 | 7.2 |
| 19 | 1 | 20 | 3.8 |
下面的公式用以计算“群体有效大小”与群体近交系数上升率之间的关系,也就是说只要进行随机交配,两者之间的关系可用下列公式计算:
△F1=1/2Ne
△F1:近交系数的上升率
Ne:群体的有效大小
由此公式我们可以看出,Ne越大,则△F1就越小。在封闭群的规定中,“群体的有效大小”被定为50。为什么要定为50与要成为近交系就必须兄弟姐妹交配20代一样,也就是说只在一般群体中,近交系数的上升率(△F1)等于百分之一时,群体的有效大小(Ne)恰好是50。
在一般情况下,大量生产实验动物的繁殖是用各种雌雄鼠的比例进行生产的。表5-3用以说明在不同性别比例时的情况,要达到“群体有效大小”50左右时所需的雌雄鼠数。根据表5-3,现假定如下计划生产方式,即每周用一只雄鼠与2只雌鼠进行交配,这种雌雄两性配合的比例为1:14。这样的群体,要达到有效大小50时,至少须经常保持雄鼠14只,雌鼠196只,这交不是说,只要准备好14只雄鼠和196只雌鼠就万事大吉了,而是为了在下一代留种时能从较多的种群中进行广泛选种,不致因种群数目较少而不得不进行近亲交配。因为,种雄雌鼠太少了,不知不觉地就提高了近交系数,结果就丧失了封闭群的意义。
表5-3雌雄性别比例不同的各群体要达到有效大小50以上时所需的雌雄鼠
| 性 别 比 例 | 群体有效大小达到50时所需的动物数 | 群体的有效大小 | |
| 雄 | 雌 | ||
| 1:1 | 25 | 25 | 50.0 |
| 1:6 | 15 | 75 | 50.0 |
| 1:7 | 15 | 106 | 52.5 |
| 1:10 | 14 | 140 | 50.9 |
| 1:14 | 14 | 196 | 52.3 |
(四)不要用人为淘汰的方式选种
所谓人为淘汰的方式选种,是把基因决定的性状引向特定方向,结果使群体内基因结构发生变化。例如在图5-3中,基因结构大体自第15代起才趋于稳定。要想增加具体a/a基因型的个体,而进行淘汰方式的选种,结果使其有a/a基因型的小鼠在全体种鼠中上升到1/2,从而使其他基因频率发生急剧变化。
图5-3所示只是一种基因型的改变所产生的结果,但即使是单基因,在实际上也伴随着许多其他遗传性状的变化。因此,在封闭群中,原则绝对上不允许进行淘汰式选种。
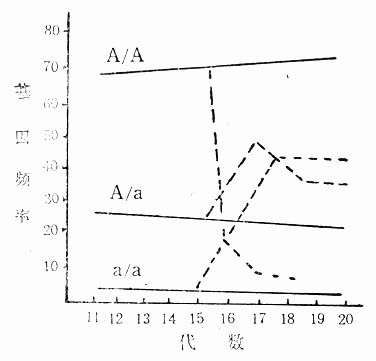
图5-3 自第15代起进行淘汰式选种后基因频率变化情况
但是,涉及到繁殖能力问题,可视为例外,因为作为实验动物首要的基本条件是可繁殖性,尤其是封闭群,利用杂合性提高繁殖率是其特特。人们在选种时有意无竟地都想用繁殖率高的动物作种。在群体被封闭后在5年之内,只要采用同一方法,只选择繁殖率高的留种,其基因结构几乎不会引起变化。因此育成封闭群后选种时,应挑选繁殖率高的为宜。
五、封闭群动物的交配方式
主要介绍来源于非近交系封闭群的交配方式。在保持封闭群时,须注意两点,第一是在群体中防止产生小群体(分化);第二是不应改变封闭群特有的杂合性。为此,决不能进行近亲交配。这里所说的近亲交配,不单是指兄弟姐妹交配。也包括父妇、母子、堂兄弟与堂姐妹间以及表亲间的交配。由于进行近亲交配,即使近交数上升,导致群体基因纯合,另一面也会丢失杂合基因,促使群体的性质发生改变。
为了防止近效系数上升,尽量增加动物的数量是最有效的措施,但群体动物的数量常受人力、物力的限制,不可能太大,而且要绝对阻止近交系数上升,目前尚缺乏有力措施,故现在只要求每代近交系数的上升率不超过1%。
为避免近交,封闭群应采取随机交配(Randommating),避免循环式的近亲交配方式。具体作法是,将群体分成若干小群,在组合下一代的交配时,有规律地把不同小群中的雌雄进行随机交配。小群体组数,可分成3、4或6,一般小群组数不宜太多,因组数太多,组合交配时比较困难,手续也比较繁杂,通常以3~4个组为宜。下图是用三组(A、B、C)小群体进行交配的方法:
由图5-4可以看到,组成新的一代时,有三种方式从A群中挑选的种雌鼠与C群中挑选的种雄鼠组成新的B群;从B小群中挑选的种雌鼠与从A群选择的种雄鼠组成新的C上述方法,以50只种雄鼠和300只种雌鼠进行繁殖,其近交系数上升率,每代不会超过0.8%左右。
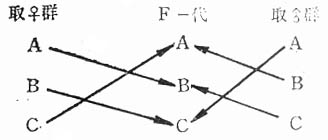
图5-4用三个群(组)保持与生产封闭群动物的示例