第二节 免疫网络学说
,。
一、Jerne的免疫网络学说
<>,、。。 <><>. -。 <>
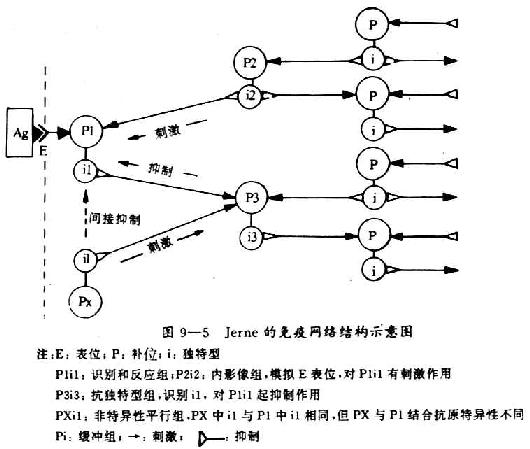 <><>.
<>():,,。()、、。,,,。,,。,,。,、。
<>():,,,,。,,。,,。
<>-。。(),,。()( )。,。( )。,,。( ),,(),,,。、、,,,。,。
<>
<><>.
<>():,,。()、、。,,,。,,。,,。,、。
<>():,,,,。,,。,,。
<>-。。(),,。()( )。,。( )。,,。( ),,(),,,。、、,,,。,。
<>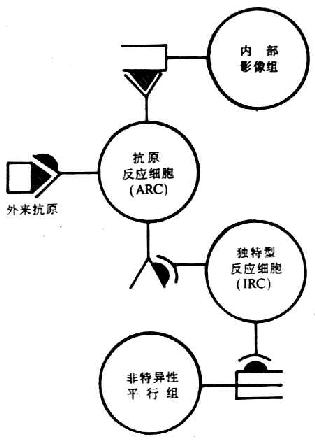 <>- ()
<>,、。
<>():β(),,。,,β。
<>():,,。,,,。,,,,,,。
<>():,。,。,,,,,,。
<>- ()
<>,、。
<>():β(),,。,,β。
<>():,,。,,,。,,,,,,。
<>():,。,。,,,,,,。
二、免疫网络的其它模型
<>、。,、。 <>,、、、,、、。,,,。,,。,,,……;,,,。,。。 <>
 <>-
<>,。
<>. ,,,,;。,,,。,,()。
<>. ,,,。
<>. ,,,,,。
<> ,()αβ, 、、[][/]。。,,α、β、。 。,,、; ,,α、β。
<>
<>-
<>,。
<>. ,,,,;。,,,。,,()。
<>. ,,,。
<>. ,,,,,。
<> ,()αβ, 、、[][/]。。,,α、β、。 。,,、; ,,α、β。
<>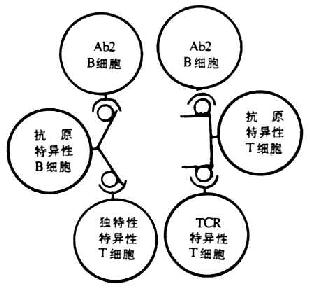 <>-
<>-
上一篇:第一节 独特型和抗独特型
下一篇:第三节 抗独特型抗体及其在医学研究中的应用