第一节 独特型和抗独特型
一、独特型发现
<>,。,()()。,,,,“”(‘')。 <>,,,。,,,。,,()(),,()(),。,,,。。,,。 <>
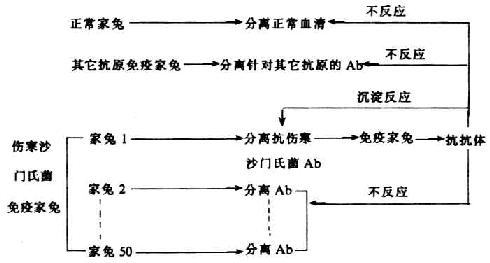 <>- (,)
<>- (,)
二、独特的分类
<>(-,α)。 <><>() <>α,:,( );,( )。 <><>() <>.() ( ,),。 <>. (- ,),。(、),。:()/,λ,,。();()。,,(-)。 <>
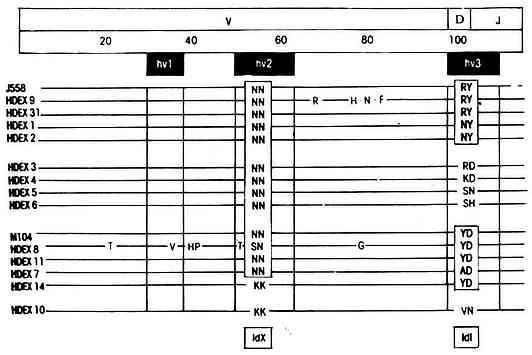 <>-
<>: : : : : :
<>: : ::
<>,()。
<>():。
<>()( ):。
<>()( ):,。。、。
<><>()
<>.()( ) ,,,。,,。
<>.( ) ,,,。
<>-
<>: : : : : :
<>: : ::
<>,()。
<>():。
<>()( ):。
<>()( ):,。。、。
<><>()
<>.()( ) ,,,。,,。
<>.( ) ,,,。
三 、独特型的分布
<>(,),(,)。,,。(,)、……,。-(-)。 <>
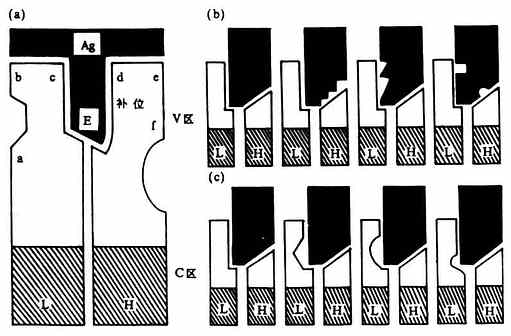 <>- 、、
<>:(),、、、、
<>()
<>()
<>,-,,。,-,。,。
<>- 、、
<>:(),、、、、
<>()
<>()
<>,-,,。,-,。,。
四、抗独特型的分类
<>,(-)。,,。 <>
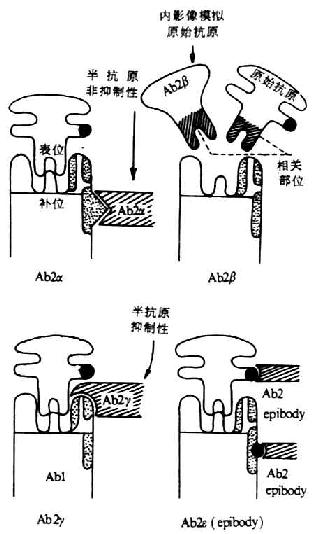 <>-
<>.α (),α,(- )。α,。,,α。
<>.β “”( ),,,β。
<>,β,β,β。,,,β。β,、、、,β,。
<>.γ (),,(- )。γ,。
<>.ε , ,,。。
<>-
<>.α (),α,(- )。α,。,,α。
<>.β “”( ),,,β。
<>,β,β,β。,,,β。β,、、、,β,。
<>.γ (),,(- )。γ,。
<>.ε , ,,。。
上一篇:第九章 免疫网络学说及其在医海陆空中的应用
下一篇:第二节 免疫网络学说