第一节 粘附分子的种类和结构
:()( );()( ,);()( );()([]+[/]- )。。
一、粘合素超家族
<>、,。,,,(),。,。 <>
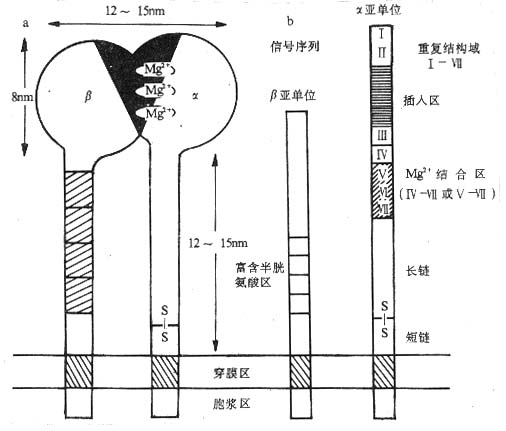 <>- ()
<>: .(),α、β ,;<>.,α([]+[/])α、 β。
<><>()
<>α、β(),α、βⅠ。α~,β~,β(β)。α(α)β(β),。αβ、、。,。。β,~;α,--------,([]+[/]),β,α 。α,,,~;,,(-)。,。
<><>()
<>αβ,αα,。αβ,αβ,αβ,α()β、β、β、ββ,βα。β,β,α。αβ(-),β、β、β、αα,。
<><>()
<>,,;,,~,。,Ⅱ/Ⅲ(Ⅱ/β);-、-、/;αβ。。
<><>()
<>。。,,,。
<>. αβ、αβ、αⅡβ、αβ、αβ、αβ,(、、、),。。
<>. αβ、αβ、αβ、αⅡβ、αβ、、、、,。,。
<> -
<><><><><><>α/β()<><><><><>< =><>(β)<>-<>/(/)<>αβ<><>,<> <><>-<>/ <>(/)<>αβ<><>,<><><>-<>+/ <>(/)<>αβ<><>,,<>?<><>-<>/ <>(/)<>αβ<><>,-<><><>-()<>+/<>(/<>αβ<><><><><>-()<>+/<>(/<>αβ<><><> <><>αβ<> <>αβ<> <><> <><>αβ<> <>αβ<> <>?<> <><>-β<>/<>(/<>αβ<> <>,<><>< =><><>(β)<>-<>/<>(/)<>αβ<><>-<>-<>-<> <><>-<>/<>(/)<>αβ<><>,<>,<>-<> <><>,<>/<>(/<>αβ<><>,<><>< => <>(β)<>ⅡⅢ<>+/<>(/)<>αⅡβ<>,,<><>,,<>,<><><><>-β<>+/<>(/)<>αβ<><>,,<>,<>,<><><>β<>αβ<>+/<>(/)<>αβ<><><> <><>β<>-β<>+/<>(/-)<>αβ<><>,<><><>β<>αβ<>+/<>(/-)<>αβ<> <><><><>β<>αβ<>(-)<>/-<>(/-)<>αβ<>αβ<> <>,-<>?<><><>β<>αβ<>/-<>(/-)<>αβ<> <>?<>
<>:(,)<>(,)<>(,)<>( ,)<>(,)<>(,)<>(,)<>( , )<>:--(--)<>:----(-----)<>:---(---)<>:---(---)<>:----(----)<>-: -,-<>-: -,-<>-: -,-<>-: -,-<>: , <>-: -,-
<>. αβ、αβ、αβ、αβ、αβ、αβ、αβ、αβ、αβ。
<><>()
<>,(,)β、β、β、ββ,、、、。
<>,α、β,。αβ,α。、、,:(),,μ/;(),。。,,。①Ⅰ( ):,;②Ⅱ( Ⅱ ):,Ⅰ;③Ⅲ( Ⅲ ):,(-)。
<>,,()Ⅲ;(),;(-)。Ⅰ、Ⅱ、Ⅲ,ⅫⅠ,Ⅱ,-Ⅲ。
<>
<>- ()
<>: .(),α、β ,;<>.,α([]+[/])α、 β。
<><>()
<>α、β(),α、βⅠ。α~,β~,β(β)。α(α)β(β),。αβ、、。,。。β,~;α,--------,([]+[/]),β,α 。α,,,~;,,(-)。,。
<><>()
<>αβ,αα,。αβ,αβ,αβ,α()β、β、β、ββ,βα。β,β,α。αβ(-),β、β、β、αα,。
<><>()
<>,,;,,~,。,Ⅱ/Ⅲ(Ⅱ/β);-、-、/;αβ。。
<><>()
<>。。,,,。
<>. αβ、αβ、αⅡβ、αβ、αβ、αβ,(、、、),。。
<>. αβ、αβ、αβ、αⅡβ、αβ、、、、,。,。
<> -
<><><><><><>α/β()<><><><><>< =><>(β)<>-<>/(/)<>αβ<><>,<> <><>-<>/ <>(/)<>αβ<><>,<><><>-<>+/ <>(/)<>αβ<><>,,<>?<><>-<>/ <>(/)<>αβ<><>,-<><><>-()<>+/<>(/<>αβ<><><><><>-()<>+/<>(/<>αβ<><><> <><>αβ<> <>αβ<> <><> <><>αβ<> <>αβ<> <>?<> <><>-β<>/<>(/<>αβ<> <>,<><>< =><><>(β)<>-<>/<>(/)<>αβ<><>-<>-<>-<> <><>-<>/<>(/)<>αβ<><>,<>,<>-<> <><>,<>/<>(/<>αβ<><>,<><>< => <>(β)<>ⅡⅢ<>+/<>(/)<>αⅡβ<>,,<><>,,<>,<><><><>-β<>+/<>(/)<>αβ<><>,,<>,<>,<><><>β<>αβ<>+/<>(/)<>αβ<><><> <><>β<>-β<>+/<>(/-)<>αβ<><>,<><><>β<>αβ<>+/<>(/-)<>αβ<> <><><><>β<>αβ<>(-)<>/-<>(/-)<>αβ<>αβ<> <>,-<>?<><><>β<>αβ<>/-<>(/-)<>αβ<> <>?<>
<>:(,)<>(,)<>(,)<>( ,)<>(,)<>(,)<>(,)<>( , )<>:--(--)<>:----(-----)<>:---(---)<>:---(---)<>:----(----)<>-: -,-<>-: -,-<>-: -,-<>-: -,-<>: , <>-: -,-
<>. αβ、αβ、αβ、αβ、αβ、αβ、αβ、αβ、αβ。
<><>()
<>,(,)β、β、β、ββ,、、、。
<>,α、β,。αβ,α。、、,:(),,μ/;(),。。,,。①Ⅰ( ):,;②Ⅱ( Ⅱ ):,Ⅰ;③Ⅲ( Ⅲ ):,(-)。
<>,,()Ⅲ;(),;(-)。Ⅰ、Ⅱ、Ⅲ,ⅫⅠ,Ⅱ,-Ⅲ。
<> <>-
<>,、、,。
<>-
<>,、、,。
二、免疫球蛋白超家族
<>、,,,( , )。。、-。。 <>、、、“”,ⅠⅡ“”。-。 <>.-( -) -,--,,-。,-,,,,,,-、-α、-;-,;-。-,,-,-~。-,,,,。,-,-(-)。--,--%,--。 <>
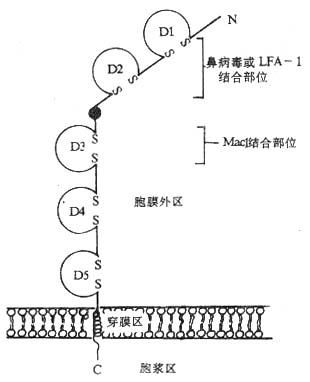 <>- -()
<>- ()、
<><><><><> <>()<> <><>-()<>,,<><>-()<><>-()<><>~<>-()<><>-()<><>~<>-()<><>-()<><><>-()<><>-()<><>/<>-()<><><>,<><>-Ⅱ()<><><>,<>/<>-Ⅰ()<><>-Ⅰ<><>/<>()<><>-Ⅱ<>,,,<>~/~<>()<><><><><>/()<><>/()<>,<><>()<><>-()<>,,<>,,<>-()<><>-()<>,,,<>,<>-()<><>-()<>,,<><>-()
<>::<>:<>:<>:<>:
<>,-。
<>。
<>.-( -),( ,),-、-α,,,--。
<>- -()
<>- ()、
<><><><><> <>()<> <><>-()<>,,<><>-()<><>-()<><>~<>-()<><>-()<><>~<>-()<><>-()<><><>-()<><>-()<><>/<>-()<><><>,<><>-Ⅱ()<><><>,<>/<>-Ⅰ()<><>-Ⅰ<><>/<>()<><>-Ⅱ<>,,,<>~/~<>()<><><><><>/()<><>/()<>,<><>()<><>-()<>,,<>,,<>-()<><>-()<>,,,<>,<>-()<><>-()<>,,<><>-()
<>::<>:<>:<>:<>:
<>,-。
<>。
<>.-( -),( ,),-、-α,,,--。
三、selectin家族
<>( ,- ).,,。 <><>() <>Ⅰ,、。,,。()()( ),,;()( - ),,,;()( )( ),(、)()。(-)。,,。 <><>() <>:-、--,、,,,。-。 <>
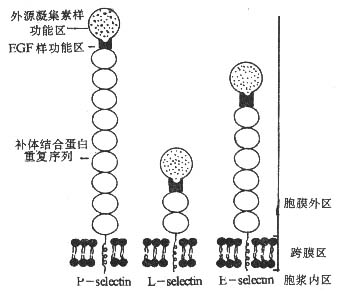 <>-
<>- 、
<><><><><><>()<><><>-(,-)<><>~<><><> ()<> <> <>-[][/]<><>-<>,<><>-[][/]<><>(,-,)<>(、、α)<> <><><>-(,-)<>(,-,)<><>-[][/]< />-[][/]< />[][/]
<>[]:[/]: ,<>: ,<>: ,<>-: -,-<>:- , <>-: -,-<>: ,
<><>()
<>,。:();();();()。。(-)(-)。,,。,,。-、- 、,、、。
<>
<>-
<>- 、
<><><><><><>()<><><>-(,-)<><>~<><><> ()<> <> <>-[][/]<><>-<>,<><>-[][/]<><>(,-,)<>(、、α)<> <><><>-(,-)<>(,-,)<><>-[][/]< />-[][/]< />[][/]
<>[]:[/]: ,<>: ,<>: ,<>-: -,-<>:- , <>-: -,-<>: ,
<><>()
<>,。:();();();()。。(-)(-)。,,。,,。-、- 、,、、。
<> <>-
<>:: : : : :
<>,,。-(---,ⅢⅡ。
<>-
<>:: : : : :
<>,,。-(---,ⅢⅡ。
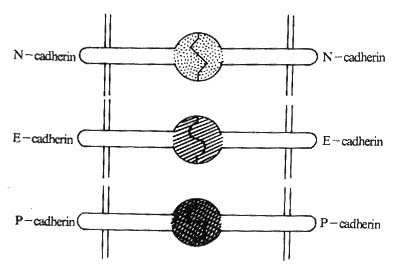
四、Cadherin家族
<>,[]+[/],,,([]+[/] )。。 <><>() <>,~,~%。Ⅰ,、。,~,,。(-)。,,,。,,,,,。 <>
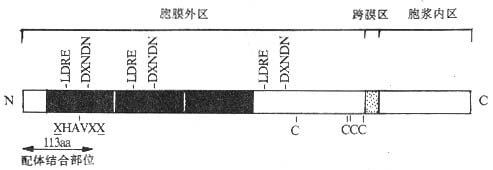 <>-
<>:;。
<><>()
<>:-、--。-、--/。,、。
<>- 、
<><><><><>()<><><><>-<><><>-<><>-<><>、、<>-<><>-<><>、、<>-
<><>()
<>,(-)。,()()。
<>
<>-
<>:;。
<><>()
<>:-、--。-、--/。,、。
<>- 、
<><><><><>()<><><><>-<><><>-<><>-<><>、、<>-<><>-<><>、、<>-
<><>()
<>,(-)。,()()。
<> <>-
<>-
五、其它未归类的粘附分子
<>,,,、-[][/]、-[][/];、、。 <><>() <>. ,。,。-[][/]-[][/],,-[][/]、,-[][/]。。 <>. ,。( ),--,~,。( ),-[][/],-。、。 <><>() <>.,-、-、Ⅲ(-Ⅲ)。,,,~、~~,。(),,,,,.,~,~。,,,-、-,。-,。-~,,,-()。。ⅠⅣ。 <>,、、、、、。 <>
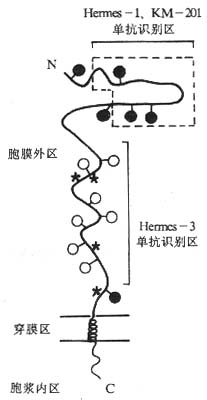 <>-
<>:●- -
<>○- ○-
<>*
<>.。
<>():,,,。:①(~),。()。( ),。②(,~),。(-),。。,。~,。,,(~)、(~)、(~)、(、)、()。
<>():,-、-。。-,-。,-,-,(-)。~,~。
<>
<>-
<>:●- -
<>○- ○-
<>*
<>.。
<>():,,,。:①(~),。()。( ),。②(,~),。(-),。。,。~,。,,(~)、(~)、(~)、(、)、()。
<>():,-、-。。-,-。,-,-,(-)。~,~。
<> <>-
<>.,-,-。
<>(),、、,。。。
<>()“”( )(),。
<>(),,/-。
<>(),。,(),。
<>()()。
<>-
<>.,-,-。
<>(),、、,。。。
<>()“”( )(),。
<>(),,/-。
<>(),。,(),。
<>()()。
上一篇:第二章 粘附分子
下一篇:第二节 粘附分子的表达的调节