第二节 淋巴细胞
参与应答的细胞(免疫细胞)可以分为三大类:第一类是指在免疫应答过程中起核心作用的免疫活性细胞,即淋巴细胞;第二类是指在免疫应答过程中起辅佐作用的单核-巨噬细胞;第三类是指单纯参与免疫效应的其他免疫细胞。由于免疫细胞的种类较多。为便于初学者掌握,将常用的细胞简称和略语列于表4-3。
淋巴细胞(lymphocyte)是受免疫系统的主要细胞,按其形成大小可分为大(11~18μm)、中(7~11μm)、小(4~7μm)三类;按其性质和功能可分为T细胞、B细胞和NK细胞。不同类型的淋巴细胞很难从形态学上分辨,只能通过其不同的表面标志和不同的反应性进行区分。
一、T细胞
T细胞是在胸腺中成熟的淋巴细胞,故称胸腺依赖性淋巴细胞(thymus-dependentlymphocyte),简称T淋巴细胞或T细胞,是血液和再循环中的主要淋巴细胞。
(一)T细胞的表面标志
在T细胞发育的不同阶段,细胞表面可表达不同种类的分子,这些分子与细胞功能有关,也可作为签别T细胞甩其活化状态的表面标志。
1.T细胞受体(Tcellreceptor,TCR)又称T细胞抗原受体,可表达于所有成熟T细胞表面,是T细胞识别外来抗原并与之结合的特异受体。TCR由α和β两个肽链组成,两肽链之间由二硫键连接,其结构和功能均类似IgG分子的一个Fab段。α链与IgG的轻链相似,由V、J和C区3个基因片段重组的基因进行编码;β链类似于IgG重链的V区和CH1区,由V、D、J和C区4个基因片段重组的基因进行编码。在T细胞发育过程中,编码α及β链的基因经历突变和重排,因此TCR具有高度的多态性,以适应千变万化的抗原分子。
与B细胞表面的Ig分子一样,TCR能特异性地与抗原结合。与Ig分子不同的是,TCR一般只结合与MHC分子连接的抗原,这是由于TCR与抗原的结合力较弱,并且常常只有α链或β链单独表达的缘故。TCR识别抗原的这种特点构成了MHC限制性的基础。TCR与抗原结合后不能直接活化T细胞,而是依赖与其邻接的CD3分子向细胞内部传递活化信息;CD4和CD8分子能够协同和加强这种作用。
一小部分成熟T细胞的表面TCR不是由α/β链组成,而是由γ/δ链组成;γ/δ链与α/β链有高度同源性,而且δ基因正好位于α基因的位点上,其意义仍然不甚明确。
2.簇分化抗原(clusterofdifferentiation,CD)T细胞在分化成熟过程中,不同的发育阶段和不同亚类的淋巴细胞可表达不同的分化抗原,这是区分淋巴细胞的重要标志。所以1986年世界卫生组织命名委员会建议应用CD系列来统一命名白细胞分化抗原,包括淋巴细胞和其它白细胞。目前已经鉴定出CD抗原70余种,部分重要CD抗原列在表4-1。
表4-1部分CD抗原及其分布和性质
| 抗原 | 别名 | 分布细胞 | 主要性质和功能 |
| CD2 | T1,E受体 | T,NK | 结合CD58,转递信号 |
| CD3 | T3,Leu4 | T | T系标志,转导信号 |
| CD4 | T4,Leu3a | T[XB]H[/XB],Mφ等EBV转化的B | MHCⅡ类分子受体,HIV受体传递信号,T[XB]H[/XB]标志 |
| CD5 | T1 | T,部分B | 结合CD72,部分B标志 |
| CD7 | Leu9 | T,NK及前体 | 传递信号 |
| CD8 | T8,Leu2a | Tc | I类分子受体,Tc标志 |
| CD10 | CALLA | B前体 | 内肽酶,急性淋巴细胞白血病的标志 |
| CD11a | LFA-1α | T,B,NK,M | 整全素,介导细胞粘附 |
| CD16 | FcγRⅢ | NK,Mφ,N | 低亲和IgGFc受体,NK标志 |
| CD19 | B4 | B | 传递信号 |
| CD21 | CR2,B2 | B | C3d、CD23和EBV受体 |
| CD22 | B | 结合CD45RO,传递信号 | |
| CD23 | FcεRⅡ | 活化B,Mφ等 | 低亲和IgEFc受体,CD21配体 |
| CD25 | IL-2Rα | 活化T,B;M | 低亲和IL-2R,L-C活化标志 |
| CD28 | 活化T[XB]H[/XB] | 结合B7,介导协同刺激信号 | |
| CD32 | FcγRⅡ | B,Mφ,N,Eo | 中亲和IgGFc受体,传递信号 |
| CD35 | CR1 | B,M,N,NK | C3b受体 |
| CD40 | B | 结合CD40,传递T[XB]H[/XB]辅助信号 | |
| CD45 | WBC-Ag | 全部WBC | PTP,传递TCR信号 |
| CD54 | ICAM-1 | 活化L-c | 结合LFA-1或CD23 |
| CD56 | N-CAM | NK | NK标志,NK粘附作用 |
| CD64 | FcγRⅠ | M,Mφ | 高亲和性IgGFc受体 |
T细胞主要的CD抗原有以下几类。
(1)CD2:分子量约49kD的糖蛋白,表达于全部人T细胞和NK细胞表面;由3种抗原性不同的分子(CD2-1,CD2-2,CD2-3)组成。CD2-1及CD2-2表达于静止细胞表面,CD2-3表达于活化的T细胞表面。应用抗CD2-2及抗CD2-3可直接活化静止的T细胞,这是成熟T细胞活化的旁路途径。CD2是粘附分子之一,顺抗原递呈过程中起辅助作用。CD2分子还可与绵羊红细胞(SRBC)结合,又称绵羊红细胞受体。在一定条件下,将外周血淋巴细胞与SRBC混合,则血T细胞能结合若干SRBC,染色后在显微镜下观察呈玫瑰花环状,故称E花环形成试验;临床上可用于测定外周血T细胞总数。
(2)CD3:为6肽复合分子,表达在全部T细胞表面,是T细胞共同的表面标志。CD3分子与TCR分子紧密连接,但CD3分子的肽链伸入胞浆的部分比TCR长得多,所以CD3可将TCR与抗原结合所产生的活化信号传递到细胞内部并激活细胞;因此应用抗CD3单克隆抗体也可直接活化T细胞。CD3与TCR的关系见图4-5。
(3)CD4/CD8:是相互关联、但意义不同的2个分子,是T细胞亚群的表面标志。表达CD4的主要是辅助性T细胞,表达CD8的主要是细胞毒性T细胞。CD4和CD8分子可增强CD3-TCR对MHC抗原的亲和力,CD4分子增强对MHCⅡ类抗原的结合,CD8分子则增强对MHCⅠ类抗原的结合。在再次免疫应答中,由于TCR-CD3与外来抗原-MHC复合分子结合的亲和力提高,即使细胞表面的CD4或CD8分子丢失,亦可发生免疫应答。
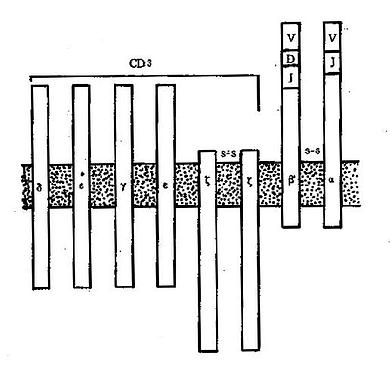
图4-5TCR与CD3结构及相关性示意图
(4)其他CD分子:CD7也是T细胞的共同标志,而且较早地出现在细胞表面,还可发现在NK细胞和少数其他淋巴样细胞前体上。某些T细胞亚群(T[XB]H[/XB])表达CD28,该分子可传递同刺激信号,与细胞活化相关。
3.其他表面标志主要包括以下几类:
(1)组织相容性抗原:T细胞主要表达MHCⅠ类抗原,个别活化的T细胞可表达MHCⅡ类抗原;MHC及其意义见第六章。
(2)致有丝分裂原受体:致有丝分裂原(mitogen)简称丝裂原,可通过相应受体刺激静止期淋巴细胞转化为淋巴母细胞,发生有丝分裂而增殖。丝裂原种类很多,常见的有植物血凝素(phytohemagglutinin,PHA)、刀豆素A(concanavalinA,ConA)等。因此可利用PHA和ConA等活化T细胞,也可借此进行淋巴细胞转化试验,判断细胞免疫的功能状态。
(3)病毒受体:淋巴细胞表面还存在病毒受体,例如麻疹病毒受体和人类免疫缺陷病毒(HIV)受体等,通过这类受体,病毒可以选择性地感染某个T细胞亚群;例如HIV可以通过CD4感染辅助性T细胞引起艾滋病。
另外,T细胞表面尚有多种白细胞介素受体(见第五章)、绵羊红细胞受体(CD2)、整合素(integrin)受体、转铁蛋白(transferrin)受体等;还有多种粘附分子,例如LFA-1和CD2等;这些均与T细胞活化有一定的相关性。
(二)T细胞的亚群及其功能
虽然T细胞有表达TCR和CD2、CD3、CD7等共性,但这是一个非均一性的复杂群体;可以分化为表达不同CD分子、具有不同免疫活性的亚群(subset),其中较为明确的两个亚群是辅助性T细胞和细胞毒性T细胞。T细胞的这种分化是不可逆的。
1.辅助性T细胞(helpTcell,T[XB]H[/XB])是能够帮助B细胞分化成抗体产生细胞和放大细胞免疫应答的一个细胞群,表达CD4但不表达CD8。T[XB]H[/XB]活化后可释放细胞因子,可以调节T细胞、B细胞、单核-巨噬细胞和其他免疫细胞的活性。T[XB]H[/XB]表面的CD4分子主要是与MHCⅡ类分子相关,而不是与辅助功能相关;因此部分CD4[SB]+[/SB]细胞有细胞毒活性,而某些CD8[SB]+[/SB]细胞有辅助作用。
根据产生细胞因子的不同,T[XB]H[/XB]可被分成3型:T[XB]H1[/XB],T[XB]H2[/XB]和T[XB]H0[/XB]。T[XB]H1[/XB]产生IL-2和IFNγ,可以增强细胞免疫应答,促进B细胞合成IgM和IgG2,活化巨噬细胞;T[XB]H2[/XB]产生IL-4和IL-5,增强IgG1和IgE的合成,增加局部和循环中嗜酸性粒细胞的数量;T[XB]H0[/XB]产生中述4类细胞因子,兼具T[XB]H1[/XB]和T[XB]H2[/XB]的生物活性。不同的抗原诱导的免疫应答可以由不同类型的T[XB]H[/XB]控制,例如蠕虫感染的免疫应答由T[XB]H2[/XB]控制,导致IgE血清水平升高和嗜酸性粒细胞数量增加;而原虫(如利什曼原虫)感染的免疫应答则由T[XB]H1[/XB]控制,产生细胞免疫应答和清除原虫的感染。
免疫应答的控制选择和T[XB]H[/XB]类型分化的机制目前尚不清楚,但细胞发育过程中的局部环境可影响细胞的分化,例如已经证明IL-4可促进T[XB]H2[/XB]应答,而IFNγ可抑制T[XB]H2[/XB]型应答。
2.细胞毒性T细胞(cytotoxicTcell,Tc或CTL)能特异性溶解靶细胞的一个细胞亚群,表达CD8但不表达CD4。Tc能够杀伤所有表达MHCⅠ类分子并向其递呈抗原的靶细胞;这种杀伤是抗原特异性的、NHCⅠ类分子限制性的和卓有成效的。Tc细胞在杀伤一个细胞后,可以转向另一个靶细胞,反复行使这种杀伤功能。因此Tc细胞在抗病毒感染、抗肿瘤免疫、移植排斥反应和某些自身免疫病中起重要作用。
3.其他T细胞亚群以往的免疫学文献曾描写过其他重要的T细胞亚群,但随着免疫学技术的进步,这些亚群很难得到证实或者已证明是别的亚群。
(1)抑制性T细胞(suppressorTcell,Ts):被描述为专门行使免疫抑制作用的T细胞亚群。当1960年首次报道某些淋巴细胞有免疫抑制作用以后,30多年来有大量的文献描述了Ts的活性,但至今没有人分离出只有免疫抑制作用的特征性T细胞群。所以目前认为:所谓抑制性T细胞,可能就是T[XB]H1[/XB],因为T[XB]H1[/XB]除了能引起细胞免疫应答外,它分泌的IFNγ还可抑制IgG1的产生。
(2)迟发型超敏反应性T细胞(delayed-typehypersensitivityTcell,T[XB]DTH[/XB]):被描述为可引起迟发型超敏反应的T细胞。有关喧个亚群的文献资料相对少些,更无人证明存在专门诱导迟发型超敏反应的T细胞群。实际上,迟发型超敏反应就是由T[XB]H[/XB]和Tc所引起的。
(3)γ/δT细胞:表面TCR由γ和δ链组成、而非通常α/β链的细胞群。这群细胞数量很少,不超过T细胞总数的5%;它们可以是CD4[SB]-[/SB]/CD8[SB]-[/SB],也可以是CD4[SB]-[/SB]/CD8[SB]+[/SB];具有细胞毒活性,也能释放细胞因子;在皮肤中存在较多,其意义有待进一步证明。
(三)T细胞的发育与分布
T细胞来源于骨髓干细胞,在胸腺内发育成熟。迁移至胸腺的初期,这些胸腺细胞只表达T细胞系的早期特征CD2和CD7,不表达CD4或CD8,称为双阴性胸腺细胞。在此期间,TCR基因开始重排;当重排的结果产生了α和β链时,TCR和CD3便开始低水平地协同表达在细胞表面。这时CD4和CD8也开始表达,并且同时出现在细胞膜上,这样的细胞称为双阳性胸腺细胞。
因双阳性胸腺细胞已经表达TCR和CD3,故能够识别与自身MHC连接的抗原;但是这时的TCR刺激不是诱导细胞增殖,而是诱导细胞凋亡(见第七章)。由于在此期间能遇到的抗原通常都是自身物质,所以死亡的细胞基本上都是自身反应细胞。这种现象称为阴性选择(negativeselection)。另一方面,能识别非自身抗原的双阳性细胞得以继续发育,其TCR和CD3分子的表达增强,并且丢失CD4或CD8分子中的一个,分化为只表达CD4或CD8的单阳性胸腺细胞,即成熟的T细胞。这种选择称为阳性选择(positiveselection)。T细胞在胸腺内的主要发育特点见图4-6。难以理解的是,阳性选择时并没有外来抗原与TCR结合,而且所选择的非自身反应性细胞却也能识别自身的MHC。
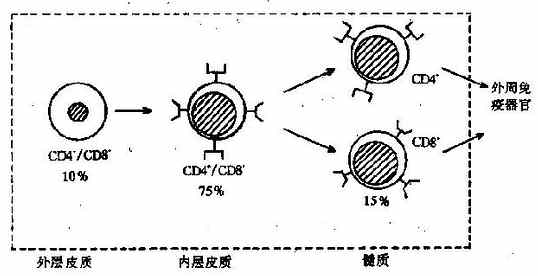
图4-6T细胞在胸腺内的发育过程
成熟的T细胞离开胸腺进入血循环,分布于外周免疫器官的胸腺依赖区,例如淋巴结的副皮质区等,受抗原刺激后参与免疫应答。T细胞是淋巴细胞再循环的主要细胞,在血液中约占60%~70%,在淋巴结约占65%~85%,在胸导管中占90%以上。当胸腺发育不全时,T细胞发育受阻,外周血淋巴细胞显著减少,外周淋巴器官的胸腺依赖区萎缩。
二、B细胞
B细胞是在鸟类法氏囊或其同功器官(骨髓)内发育成熟的细胞,因此称为法氏囊或骨髓依赖的淋巴细胞(bursaorbonemarrowdependentlymphocyte),简称B淋巴细胞或B细胞。B细胞受抗原刺激后可产生抗体的浆细胞。
(一)B细胞的表面标志
1.表面免疫球蛋白(surfaceimmunoglobulin,SLg)是B细胞最具特征性的表面标志。成熟B细胞膜表面表达SIgM和SigD,早期B细胞只表达SIgM分子;SIg的肽链结构与Ig相同,但SIgM是单体分子。一个B细胞表面可有上万个SIg分子,其特异性都与细胞分泌的Ig分子相同。由于细胞在分化过程中基因突变和Ig基因重排的结果,一个正常人体内至少有3×10[SB]6[/SB]个可产生不同抗体分子的细胞克隆。SIg的功能是为B细胞表面的抗原受体,可与相应抗原特异性结合,并将抗原作内摄处理。这种受体介导的结合是B细胞捕获抗原的主要方式。
SIgM或SIgD分子的羧基端插入细胞膜,但只有几个氨基酸的深度;所以象TCR一样,SIg自身不能独立地向细胞内传递刺激信号,必须依赖与其紧密相关的其他两种跨膜糖蛋白棗Ig-α和Ig-β。这两种蛋白靠二硫键连在一起,各有一个大的肽段伸入细胞浆,因此可以象T细胞表面的CD3协同TCR一样辅助SIg向细胞内传递刺激信号。
2.MHC和CD抗原成熟的B细胞表面表达MHCⅡ类分子,这可使B细胞作为抗原递呈细胞,与其免疫活性相关(详见第七章)。
B细胞在分化成熟过程中可表达不同的CD分子,其中某些可作为B细胞的标志,某些还与细胞功能相关。CD10只出现在B前体细胞,CD19从原始至成熟的B细胞都存在,而CD22只在成熟B细胞表达,都是经常检测的标志;CD21和CD35是补体受体;CD23和CD32则是Ig的Fc受体;CD40可与T[XB]H[/XB]细胞上的配体相结合,从而接受T[XB]H[/XB]的辅助作用。
3.其他标志及作用
(1)Fc受体:B细胞表面有IgG的Fc受体(CD32),与B细胞活性有关。Fc受体还可与抗体包被的红细胞相结合形成EAC玫瑰花环,是鉴别B细胞的传统方法之一。
(2)补体受体:CR表达于成熟B细胞的表面,CR1可与C3b和C4b结合,促进B细胞活化或抑制补体活化;CR2受体(CD21)可与C3d结合,同时也是EB病毒的受体。
(3)丝裂受体:B细胞的致有丝分裂原主要是脂多糖(LPS),受丝裂原活化后B细胞也可以分化增殖。
另外,B细胞表面还有多种细胞因子(如IL-1,IL-2,IL-4,和IFNγ等)的受体,与不同细胞因子的结合可使B细胞产生相应的生物活性。B细胞与T细胞的比较见表4-2。
表4-2T细胞、B细胞与NK细胞的性状比较
| 性状 | T细胞 | B细胞 | NK细胞 |
| 分化成熟部位 | 胸腺 | 骨髓 | ? |
| 表面标志 | |||
| 表面膜Ig | - | + | - |
| TCR | + | - | - |
| HLAⅡ类抗原 | 极少数 | + | - |
| CD2 | + | - | + |
| CD3 | + | - | - |
| CD19,CD20 | - | + | - |
| CD16,CD56 | - | - | + |
| CR | - | + | 部分 |
| FCγR | - | + | + |
| 细胞分布(%) | |||
| 外周血 | 60~70 | 20~30 | 10~15 |
| 骨髓 | <5 | >95 | 极少 |
| 胸导管 | 90 | 10 | 少见 |
| 脾 | 30~50 | 50~60 | 少见 |
| 淋巴结 | 65~85 | 15~35 | 少见 |
(二)B细胞的分化成熟
人和哺乳动物B细胞的产生、发育和成熟均在骨髓中完成,这个过程伴随着一系列的胞内基因和表面标志的变化(图4-7)。
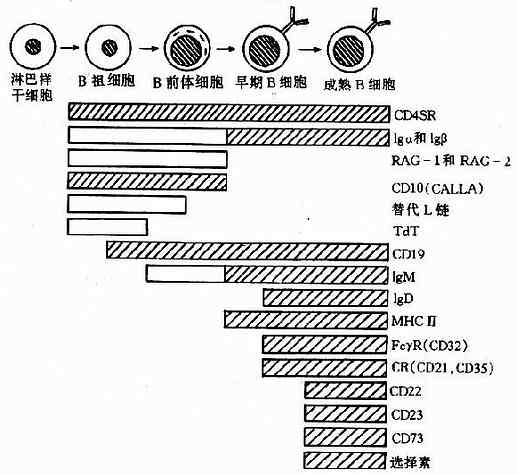
图4-7B细胞发育过程及表面特征
B祖细胞(pro-Bcell)只表达CD10、CD19的末端脱氧核苷转移酶(TdT),这个时期的主要变化是重链基因重排;重排的失败率占50%,不成功的重排导致细胞死亡。成功的重排可导致产生重链μm,存放于粗面内质网中;重链μm可与未重排轻链基因产物化替轻链(surrogatelightchains)相结合,所形成的临时复合物能转移到细胞膜上;这种转移产生的信号使重链基因重排停止,表面临时复合物消失,细胞发育进入下一阶段。
B前体细胞(pre-Bcell)在进入成熟前不再分裂增殖,这时期的主要变化是轻链基因重排。重排的基因产生κ或λ链,与已有的重链结合形成4肽的IgM单体,并转移到细胞膜上。这一信号使细胞进入成熟期,永久性地失去Ig基因重排的能力;因此受抗原刺激后增殖的所有分子代细胞均产生与SIg同样特异性的抗体分子。成熟B细胞还表达其他表面标志。例如IgD、归巢受体、MHCⅡ类分子、CD22、CD23和CD40等。
(三)B细胞的亚群和分布
不同克隆的成熟B细胞表达不同特异性的抗原受体,自然地体现了它们之间的差别,但是很难找到其他的显著标志(例如象T细胞的CD4和CD8那样)将B细胞分成几个亚群。近年来发现一小部分成熟B细胞表达CD5,这些细胞多与自身反应(产生自身抗体)相关;CD5还发现在几乎所有慢性淋巴性白血病细胞上。
未成熟B细胞都在骨髓内,成熟初期仍留在骨髓2~3天,待表面标志表达完全后便离开骨髓进入血循环,分布到外周免疫器官的非胸腺依赖区。B细胞在血液中约占20%~30%,在胸导管中不超过10%,在淋巴结中约占15%~35%,在脾中数量最多,可达60%。B细胞受抗原刺激后可在外周免疫器官中继续增殖分化为浆细胞,分泌抗体(详见第七章)。
三、自然杀伤细胞
自然杀伤细胞(naturalkiller,NK)可非特异性杀伤肿瘤细胞和病毒感染细胞的淋巴样细胞。目前对NK细胞的了解尚不甚清楚,但其免疫学潜能值得进一步探讨。
(一)NK细胞的一般特征
NK细胞形体较大,含有胞浆颗粒,在形态上独具特色。NK细胞表面没有TCR和CD3等T细胞标志,也没有SIg和CD40等B细胞标志,因此曾被称为无标志细胞或裸细胞(nullcell)。NK细胞表达CD56,可作为细胞的系统标志;与单核-巨噬细胞一样表达CD16等分子,与其杀伤功能相关。
NK细胞也来源于骨髓前体细胞,其发育地点和分化过程尚不明确;但其发育环境和分化显然不同于T细胞和B细胞,因为T和B细胞联合免疫缺陷时NK细胞可正常;而T和B细胞正常时可有NK细胞缺陷。
NK细胞的数量较少,在外周血中约占淋巴细胞总数的15%,在脾中约3%~4%,也可发现在肺、肝和肠粘膜;但在胸腺、淋巴结和胸导管中罕见。
(二)NK细胞的活性和功能
1.自然杀伤活性NK细胞的主要活性是杀伤肿瘤细胞和病毒感染细胞;与Tc细胞不同,这种杀伤不需要TCR识别靶细胞上的抗原,也不需要识别靶细胞上的MHC分子,因此可以在靶细胞暴露的早期行使杀伤功能,不需要事先的抗原致敏,所以称为自然杀伤。
自然杀伤的识别机制尚不清楚。一种重要的观点是NK细胞表面可表达两型受体:一为刺激型,可传递信号使细胞释放IFNγ和THFα等细胞因子,从而杀伤细胞;另一为相反作用的抑制型。当NK细胞只表达刺激型受体时才有杀伤作用。现已知抑制型受体是一种称为Ly-49的膜表面分子,它的配体是MHCⅠ类分子;当细胞表达Ⅰ类分子旺盛时,NK细胞便失去自然杀伤的能力。
2.细胞因子活化的杀伤作用NK细胞的杀伤活性可通过某些细胞因子(例如IL-2)的诱导而显著增强,这样的细胞称为淋巴因子活化的杀伤细胞(lymphokine-activatedkiller,LAK)。外周血单个核细胞在含IL-2的培养基中孵育便可得到LAK细胞,NK细胞是其主要成分。用LAK细胞治疗肿瘤是颇有潜力的一种生物疗法。
3.抗体依赖的杀伤作用NK细胞表面有IgG的Fc受体(CD16),因此亦可通过抗体的媒介活化NK细胞,杀伤抗体包被的靶细胞,这种特殊的活性称为抗体依赖性细胞介导的细胞毒作用(antibody-dependentcell-mediatedcytotoxicity,ADCC)。在这种情况下,对靶细胞的识别选择取决于特异性抗体。