第三节 休克的病理生理变化
一、微循环变化
各种休克虽然由于致休克的动因不同,在各自发生发展过程中各有特点,但微循环障碍(缺血、淤血、播散性血管内凝血)致微循环动脉血灌流不足,重要的生命器官因缺氧而发生功能和代谢障碍,是它们的共同规律。休克时微循环的变化,大致可分为三期,即微循环缺血期、微循环淤血期和微循环凝血期。下面以低血容量性休克为例阐述微循环障碍的发展过程及其发生机理。
低血容量性休克常见于大出血、严重的创伤、烧伤和脱水。其微循环变化发展过程比较典型(图10-1)。
(一)微循环缺血期(缺血性缺氧期)
此期微循环变化的特点是:①微动脉、后微动脉和毛细血管前括约肌收缩,微循环灌流量急剧减少,压力降低;②微静脉和小静脉对儿茶酚胺敏感性较低,收缩较轻;③动静脉吻合支可能有不同程度的开放,血液从微动脉经动静脉吻合支直接流入小静脉。
引起微循环缺血的关键性变化是交感神经——肾上腺髓质系统强烈兴奋。不同类型的休克可以通过不同机制引起交感——肾上腺髓质性休克和心源性休克时,心输出量减少和动脉血压降低可通过窦弓反射使交感——肾上腺髓质系统兴奋;在大多数内毒素性休克时,内毒素可直接剌激交感——肾上腺髓质系统使之发生强烈兴奋。
交感神经兴奋、儿茶酚胺释放增加对心血管系统的总的效应是使外周总阻力增高和心输出量增加。但是不同器官血管的反应却有很大的差别。皮肤、腹腔内脏和肾的血管,由于具有丰富的交感缩血管纤维支配,。而且α受体又占有优势,因而在交感神经兴奋、儿茶酚胺增多时,这些部位的小动脉、小静脉、微动脉和毛细血管前括红肌都发生收缩,其中由于微动脉的交感缩血管纤维分布最密,毛细血管前括约肌对儿茶酚胺的反应性最强,因此它们收缩最为强烈。结果是毛细血管前阻力明显升高,微循环灌流量急剧减少,毛细血管的平均血压明显降低,只有少量血液经直捷通路和少数真毛细血管流入微静脉、小静脉,组织因而发生严重的缺血性缺氧。脑血管的交感缩血管纤维分布最少,α受体密度也低,口径可无明显变化。冠状动脉虽然也有交感神经支配,也有α和β受体,但交感神经兴奋和儿茶酚胺增多却可通过心脏活动加强,代谢水平提高以致扩血管代谢产物特别是腺苷的增多而使冠状动脉扩张。
交感兴奋和血容量的减少还可激活肾素-血管紧张素-醛固酮系统,而血管紧张素Ⅱ有较强的缩血管作用,包括对冠状动脉的收缩作用。
此外,增多的儿茶酚胺还能剌激血小板产生更多的血栓素A[XB]2[/XB](thromboxane A[XB]2[/XB],TXA[XB]2[/XB]),而。TXA[XB]2[/XB]也有强烈的缩血管作用。
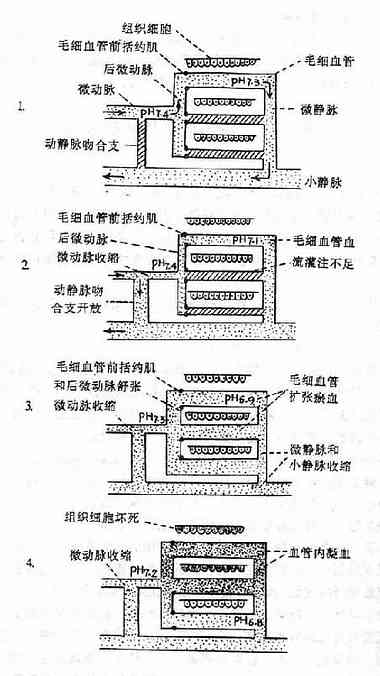
图10-1 微循环障碍的发展过程模式图
1.正常情况
⑴动静脉吻合支是关闭的。
⑵只有20%毛细血管轮流开放,有血液灌流。
⑶毛细血管开放与关闭受毛细血管前括约肌的舒张与收缩的调节。
2.微循环缺血期
⑴交感神经兴奋和肾上腺素、去甲肾上腺素分泌增多,小动脉、微动脉、后微动脉,毛细血管前括约肌收缩。
⑵动静脉吻合支开放,血液由微动脉直接流入小静脉。
⑶毛细血管血液灌流不足,组织缺氧。
3.微循环淤血期
⑴小动脉和微动脉收缩,动静脉吻合支仍处于开放状态,进入毛细血管的血液仍很少。
⑵由于组织缺氧,组织胺、缓激肽、氢离子等舒血管物质增多,后微动脉和毛细血管前括约肌舒张,毛细血管开放,血管容积扩大,进入毛细血管内的血液流动很慢。
⑶由于交感神经兴奋,肾上腺素和去甲肾上腺素分泌增多(可能还有组织胺的作用),使微静脉和小静脉收缩,毛细血管后阻力增加,结果毛细血管扩张淤血。
4.微循环凝血期
⑴由于组织严重缺氧、酸中毒,毛细血管壁受损害和通透性升高,毛细血管内血液浓缩,血流淤滞;另外血凝固性升高,结果在微循环内产生播散性血管内凝血。
⑵由于微血栓形成,更加重组织缺氧和代谢障碍,细胞内溶酶体破裂,组织细胞坏死,引起各器官严重功能障碍。
⑶由于凝血,凝血因子(如凝血酶原、纤维蛋白原等)和血小板大量被消耗,纤维蛋白降解产物增多,又使血液凝固性降低;血管壁又受损害,继而发生广泛性出血。
而TXA[XB]2[/XB]也有强烈的缩血管作用。
还有,溶酶体水解酶-心肌抑制因子系统在休克Ⅰ期微循环缺血的发生中也起一定的作用。休克时,主要由于胰腺血液灌流量减少所引起的缺血、缺氧和酸中毒可使胰腺外分泌细胞的溶酶体破裂而释出组织蛋白酶,后者即可分解组织蛋白而生成心肌抑制因子(myocardial depressant factor, MDF)。小分子肽MDF进入血流后,除了引起心肌收缩力减弱、抑制单核吞噬细胞系统的吞噬功能以外,还能使腹腔内脏的小血管收缩,从而进一步加重这些部位微循环的缺血。
本期的主要临床表现是:皮肤苍白,四肢厥冷,出冷汗,尿量减少;因为外周阻力增加,收缩压可以没有明显降低,而舒张压有所升高,脉压减小,脉搏细速;神志清楚,烦躁不安等。
此期微循环变化具有一定的代偿意义。皮肤和腹腔器官等小动脉收缩,既可增加外周阻力,以维持血压,又可减少这些组织器官的血流量,以保证心脑等重要器官的血液供给;毛细血管前阻力增加,毛细血管流体静压降低,促使组织液进入血管,以增加血浆容量;另外,动静脉吻合支开放,静脉收缩使静脉容量缩小(正常约有70%血液在静脉内),可以加快和增加回心血量,也有利于血压的维持和心脑的血液供给。但是由于大部分组织器官因微循环动脉血灌流不足而发生缺氧,将导致休克进一步发展。如能及早发现,积极抢救,及时补充血量,降低过剧的应激反应,可以很快改善微循环和恢复血压,阻止休克进一步恶化,而转危为安。
这时微循环变化的机理可概括如下(图10-2):
(二)微循环淤血期(淤血性缺氧期)
在休克的循循环缺血期,如未能及早进行抢救,改善微循环,则因组织持续而严重的缺氧,而使局部舒血管物质(如组织胺、激肽、乳酸、腺苷等)增多,后微动脉和毛细血管前括约肌舒张,微循环容量扩大,淤血,发展为休克微循环淤血期。此期微循环变化的特点是:①后微动脉和毛细血管前括约肌舒张(因局部酸中毒,对儿茶酚胺反应性降低),毛细血管大量开放,有的呈不规侧囊形扩张(微血池形成),而使微循环容积扩大;②微静脉和小静脉对局部酸中毒耐受性较大,儿茶酚胺仍能使其收缩(组织胺还能使肝、肺等微静脉和小静脉收缩),毛细血管后阻力增加,而使微循环血流缓慢;③微血管壁通透性升高,血浆渗出,血流淤滞;④由于血液浓缩,血细胞压积增大,红细胞聚集,白细胞嵌塞,血小板粘附和聚集等血液流变学的改变,可使微循环血流变慢甚至停止。⑤由于微循环淤血,压力升高,进入微循环的动脉血更少(此时小动脉和微动脉因交感神经作用仍处于收缩状态)。由于大量血液淤积在微循环内,回心血量减少,使心输出量进一步降低,加重休克的发展。
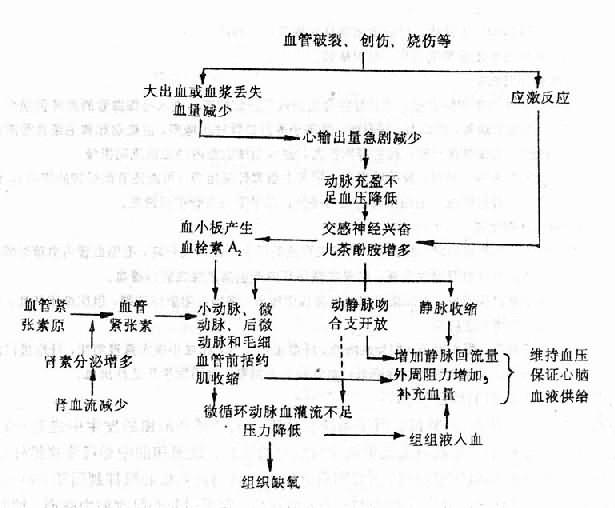
图10-2 缺血性缺氧期微循环变化机理
由于上述微循环变化,虽然微循环内积有大量血液,但动脉血灌流量将更加减少,病人皮肤颜色由苍白而逐渐发绀,特别是口辰和指端。因为静脉回流量和心输出量更加减少,病人静脉萎陷,充盈缓慢;动脉压明显降低,脉压小,脉细速;心脑因血液供给不足,ATP生成减少,而表现为心收缩力减弱(心音低),表情淡漠或神志不清。严重的可发生心、肾、肺功能衰竭。这是休克的危急状态,应立即抢救,补液,解除小血管痉挛,给氧,纠正酸中毒,以疏通微循环和防止播散性血管内凝血。这时微循环变化的机理可概括如下(图10-3):
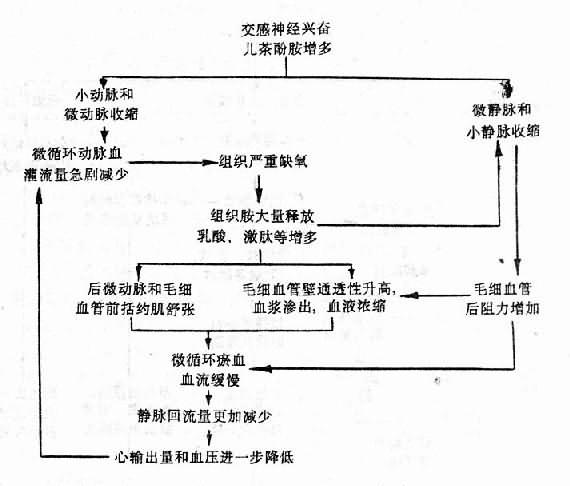
图10-3 淤血性缺氧期微循环变化机理
(三)微循环凝血期(播散性血管内凝血)
从微循环的淤血期发展为微循环凝血期是休克恶化的表现。其特点是:在微循环淤血的基础上,于微循环内(特别是毛细血管静脉端、微静脉、小静脉)有纤维蛋白性血栓形成,并常有局灶性或弥漫性出血;组织细胞因严重缺氧而发生变性坏死。
播散性血管内凝血与休克的联系极为密切。关于播散性血管内凝血引起的病理变化以及它如何引起休克或加重休克的发展,已在《播散性血管内凝血》一章讨论过了,这里再概要地归纳一下休克如何引起播散性血管内凝血。
1.应激反应使血液凝固性升高。致休克的动因(如创伤、烧伤、出血等)和休克本身都是一种强烈的剌激,可引起应激反应,交感神经兴奋和垂体-肾上腺皮质活动加强,使血液内血小板和凝血因子增加,血小板粘附和聚集能力加强,为凝血提供必要的物质基础。
2.凝血因子的释放和激活。有的致休克动因(如创伤、烧伤等)本身就能使凝血因子释放和激活。例如,受损伤的组织可释放出大量的组织凝血活素,起动外源性凝血过程;大面积烧伤使大量红细胞破坏,红细胞膜内的磷脂和红细胞破坏释出的ADP,促进凝血过程。
3.微循环障碍,组织缺氧,局部组织胺、激肽、乳酸等增多。这些物质一方面引起毛细血管扩张淤血,通透性升高,血流缓慢,血液浓缩红细胞粘滞性增加,有利于血栓形成;另一方面损害毛细血管内皮细胞,暴露胶元,激活凝血因子Ⅻ和使血小板粘附与聚集。
4.缺氧使单核吞噬细胞系统功能降低,不能及时清除凝血酶元酶、凝血酶和纤维蛋白。结果在上述因素作用下,而发生播散性血管内凝血(图10-4)。
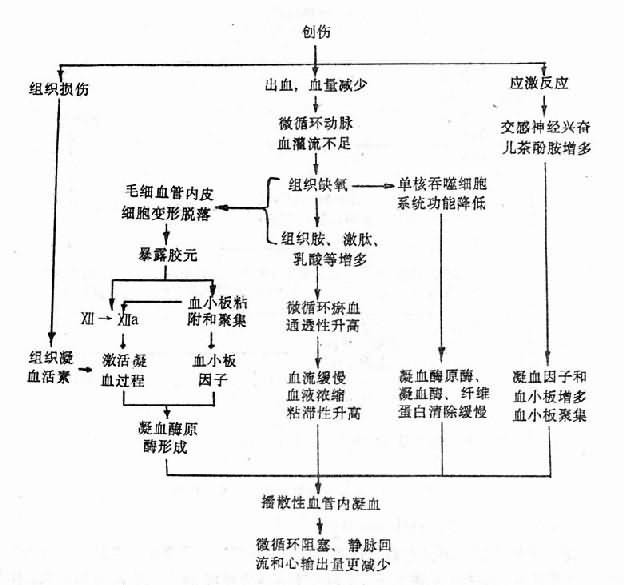
图10-4 创伤性休克引起播散性血管内凝血的机理
应当指出,在不同类型的休克,播散性血管内凝血形成的早晚可不相同。例如,在烧伤性和创伤性休克时,由于有大量的组织破坏,感染中毒性休克时,由于内毒素对血管内皮的直接损伤,因而都可较早地发生播散性血管内凝血,而在失血性休克等,则播散性血管内凝血发生较晚。
播散性血管内凝血一旦发生,将使微循环障碍更加严重,休克病情进一步恶化,这是因为:①广泛的微血管阻塞进一步加重微循环障碍,使回心血量进一步减少;②凝血物质消耗、继发纤溶的激活等因素引起出血,从而使血容量减少;③可溶性纤维蛋白多聚体和其裂解产物等都能封闭单核吞噬细胞系统,因而使来自肠道的内毒素不能被充分清除。
由于播散性血管内凝血的发生和微循环淤血的不断加重,由于血压降低所致的全身微循环灌流量的严重不足,全身性的缺氧和酸中毒也将愈益严重;严重的酸中毒又可使细胞内的溶酶体膜破裂,释出的溶酶体酶(如蛋白水解酶等)和某些休克动因(如内毒素等)都可使细胞发生严重的乃至不可逆的损害,从而使包括心、脑在内的各重要器官的机能代谢障碍也更加严重(详后),这样就给治疗造成极大的困难,故本期又称休克难治期。
二、血液流变学的变化
血液流变学(hemorheology)是研究血液流动和变形的科学,或者说是研究血液的流变性、凝固性、血液有形成分(主要是红细胞)粘弹性以及心血管的粘弹性和变形的科学。物体在一定外力作用下能流动或变形的特性,称为该物体流变性。一切流体在一定外力作用下,都具有流动性,但流动的难易,则主要取决于流体内部对于流动起阻抗作用的分子之间和颗粒之间的内摩擦力(即流体的粘度)。例如,水的粘度低,容易流动,即流度大;血液的粘度大(红为蒸馏水的4-5倍),不易流动,即流度小。由于流体的流动是以物体的变形为基础,所以流体的粘度是映流体流变性的重要指标。
血液是由水、无机盐、蛋白质、脂类、糖等大小分子所组成的混合液,其中还悬浮着大量具有可塑性的红细胞,所以血液是一种高浓度的悬浊液。因此能够影响血液流变性的因素主要有:血细胞压积(血液粘度随血细胞的压积增加而升高)、血细胞的分散程度(血细胞处于分散状态,血液粘度较低;红细胞或血小板发生聚集,血液粘度升高)、红细胞的可塑性(红细胞可塑性降低,不易变形,血液粘度增加)、血浆内高分子化合物的浓度(血浆粘度大小与其所含蛋白质、脂类、糖等的浓度呈正比)、血管内壁平滑度(血管内皮受损、变形,流经的血液粘度升高)。此外,与血管的长度、口径、血管壁的弹性和张力也有关系。
休克时血液流变学的主要变化是:
1.血细胞比容血细胞比容的改变与休克的原因和发展阶段有关。在低血容量性休克的早期,由于组织间液向血管内转移,导致血液稀释,血细胞比容降低,当休克进入微循环淤血期,由于微血管内流体静压升高和毛细血管通透性增高,液体乃从毛细血管内外渗至组织间隙,因而血液浓缩,血细胞比容升高。血细胞比容越高,血液粘度越大,血流阻力越大,而血流量则越少,血流更加缓慢。
2.红细胞变形能力降低,聚集力加强在正常情况下,红细胞在流经小于其直径的毛细血管时,可折叠、弯曲而发生多种变形以减少其宽度,从而得以顺利通过。现已证明,休克时红细胞的变形能力明显降低,其主要原因是:①休克Ⅱ期时因血液浓缩和组织缺氧所引起的血液渗透压升高和pH降低,可使红细胞膜的流动性和可塑性降低并使红细胞内部的粘度增加;②ATP缺乏(可由缺氧或某些休克动因直接引起)可使红细胞不能维持正常的功能和结构。结果是由于红细胞的变形能力降低而难以通过毛细血管,从而导致血流阻力增高。
红细胞聚集加强,是休克时细胞流变学的重要改变之一。轻者4、5个红细胞聚集在一起,重者20~30个红细胞聚集成长链或团块。引起红细胞聚集的原因是:①血流速度变慢,切变率(shear rate)降低:正常时由于血流速度快和切变率高。可防止红细胞的聚集,并可促使聚集的红细胞解聚。休克时随着血压下降,血液流速减慢和切变率降低,红细胞就易于聚集。②红细胞表面电荷减少:正常红细胞表面因含有唾液酸的羧基,故都带有负电荷。红细胞之间的这种同电荷的排斥力可阻止红细胞互相靠拢和聚集。休克时,尤其是内毒素性休克时,红细胞表面负电荷减少,可能是由于血浆中带正电荷的蛋白质增多,被红细胞吸附所致,从而使红细胞彼此靠拢发生聚集。③血细胞比容增加:已如前述,休克时由于血浆外渗,血液浓缩,故血细胞比容增加,这就可以促进红细胞聚集。④纤维蛋白原浓度增高;纤维蛋白原覆盖于红细胞表面,在红细胞之间可形成有相互聚集作用的“桥力”。休克时由于纤维蛋白原浓度增高,致使“桥力”增大乃至超过负电荷的排斥力。因而就可导致红细胞的聚集。红细胞聚集轻则增加血液粘度和血流阻力,重则可引起红细胞淤滞并阻塞微循环,甚至形成微血栓。
(一)白细胞粘着和嵌塞
正常微循环的血流是红细胞位于中央的轴流,血浆构成边流,虽然也可见到少量白细胞附壁滚动,但不发生附壁粘着现象。休克时可见白细胞附着于小静脉壁,致使血流阻力增高和静脉回流障碍。发生白细胞附壁的原因可能与白细胞和管壁之间吸引力增大,休克时血流变慢和切应力(shear stress)下降等因素有关。休克时,还可见到白细胞嵌塞于血管内皮细胞核的隆起处或毛细血管分支处,这可增加血流阻力和加重微循环障碍,而且嵌塞的白细胞还可释放自由基和溶酶体酶类物质,从而破坏生物膜和引起坏死。休克时白细胞发生嵌塞的原因是:①白细胞的变形能力降低,故不易通过毛细血管而发生嵌塞;②休克时血压下降,脉压差减小,动脉血流量减少,驱动白细胞通过毛细血管的力量减弱,因而易于发生白细胞嵌塞。
(二)血小板粘附和聚集
血小板粘附是指血小板和血小板以外的物质相互粘附的现象,血小板聚集则是血小板之间相互发生反应并形成血小板团(或称血小板聚集物)的过程。粘附一旦开始,聚集过程也随之发生。在血小板聚集开始时,其表面首先失去光滑性,变得粗糙,形成有突剌的球状体(或称聚集型血小板)。在内毒素性、创伤性和烧伤性休克时,血液中这种聚集型血小板的数目增多,而且在微血管中有血小板粘附、聚集和血小板微血栓的形成。这种聚集的血小板不但阻塞微血管,还可释放多种生物活性物质如儿茶酚胺、TXA[XB]2[/XB]、5-羟色胺等,使局部微血管收缩、通透性增高、血管内皮水肿和血流减少。此外,尚可释放促凝血的血小板因子(如PF[XB]3[/XB]等),加速凝血过程,形成DIC。
休克时引起血小板粘附和聚集的主要原因是:①血流减慢,血管内皮完整性破坏,内膜下胶原暴露,为血小板粘附提供了基础;②损伤的内皮组织释放ADP,发生聚集的血小板可释放ADP、TXA[XB]2[/XB]以及血小活化因子(PAF),均可触发并加重血小板的聚集。
(三)血浆粘度增大
休克时,尤其是严重创伤或烧伤休克时,一方面由于机体发生应激,使体内合成纤维蛋白原增多;另一方面,在休克的微循环淤血期,毛细血管内的流体静压增高,微血管周围的肥大细胞又因缺氧而释放组胺并从而使毛细血管通透性增高,液体乃从毛细血管大量外渗至组织间隙,因而血液浓缩,使血浆纤维蛋白原浓度增高,有时纤维蛋白原可高达10g/L(1000mg/dl),故可使血浆粘度增大。这不但影响组织血液流量,并可促进红细胞的聚集。如当纤维蛋白原的浓度增到5~8g/L(500~800mg/dl)时,由于血浆粘度的增高,红细胞就发生聚集,形成缗钱状。
总之,由于发生上述血液流变学的改变,不但会加重微循环障碍和组织的缺血缺氧,还可促进DIC的形成和休克的发展,近年来应用血液稀释治疗休克,其目的在于改善血液流变学,降低血流粘度。这种疗法已取得良好的效果。
三、细胞代谢的变化以及功能、结构的损害
休克时细胞的代谢障碍及其功能、结构的损害,既是组织低灌流、微循环流变学改变和/或各种毒性物质作用的结果,又是引起各重要器官功能衰竭和导致不可逆性休克的原因。
(一)休克时细胞的代谢变化
休克时细胞代谢改变比较复杂。由于休克的类型、发展价段以及组织器官的不同,其代谢改变的特点和程度也都有所不同,但共同的重要改变是:
1.糖酵解加强休克时由于组织的低灌流和细胞供氧减少,使有氧氧化受阻,无氧酵解过程加强,从而使乳酸产生增多,而导致酸中毒。但严重酸中毒又可抑制糖酵解限速酶如磷酸果糖激酶等的活性,使糖酵解从加强转入抑制。
2.脂肪代谢障碍正常情况下,脂肪分解代谢中产生的脂肪酸随血液进入细胞浆后,在脂肪酰辅酶A(脂肪酰CoA)合成酶的作用和ATP的参与下,被活化为水溶性较高的的脂肪酰CoA,后者再经线粒体膜上肉毒碱脂肪酰转移酶的作用而进入线粒体中,通过β-氧化生成乙酰辅酶A,最后进入三羧酸循环被彻底氧化。休克时,由于组织细胞的缺血缺氧和酸中毒,使脂肪酰CoA合成酶和肉毒碱脂肪酰转移的活性降低,因而脂肪酸的活化和转移发生障碍;另方面因线粒体获氧不足和/或某些休克动因(如细菌内毒素)、酸中毒等的直接作用使线粒体呼吸功能被抑制,使转入线粒体内的脂肪酰CoA不能被氧化分解,结果造成脂肪酸和/或脂肪酰CoA在细胞内蓄积,从而加重细胞的损害。
(二)休克时细胞的损害
休克时细胞的损害首先是生物膜(包括细胞膜、线粒体膜和溶酶体膜等)发生损害。
1.细胞膜的损害 最早的改变是细胞膜通透性增高,从而使细胞内的Na[SB]+[/SB]、水含量增加而K[SB]+[/SB]则向细胞外释出,细胞膜内外Na[SB]+[/SB]、K[SB]+[/SB]分布的变化,使细胞膜Na[SB]+[/SB]-K[SB]+[/SB]ATP酶活性增高。因而ATP消耗增加,再加上ATP的供应不足和膜上受体腺苷酸环化酶系统受损,结果使控制细胞代谢过程的第二信使-cAMP含量减少,因此细胞的许多代谢过程发生紊乱,例如休克时肌肉细胞对胰岛素的反应减弱,使胰岛素促进细胞摄取葡萄糖的效应减弱甚至丧失。
休克时引起细胞膜损害的原因是多方面的:
(1)能量代谢障碍休克时因组织细胞的缺血缺氧,一方面ATP生成不足,使细胞膜不能维持正常功能和结构;另一方面脂肪酸氧化受阻,蓄积于细胞内的脂肪酸和脂肪酰CoA与细胞内Na[SB]+[/SB]、K[SB]+[/SB]、Ca[SB]+[/SB]等阳性离子结合形成“皂类”化合物,可直接对膜上脂类起“净化去垢”的破坏作用。
(2)细胞酸中毒休克时细胞发生酸中毒,除与乳酸等蓄积有关外,还可能与下述因素有关:①细胞低灌流,使产生的CO[XB]2[/XB]不易排出;②ATP分解过程中产生H[SB]+[/SB](MgATP[SB]2-[/SB]→MgADP[SB]-[/SB]+Pi[SB]2-[/SB]+H[SB]+[/SB]);③胞浆Ca[SB]2+[/SB]增多,可促使Ca[SB]2+[/SB]进入线粒体并与其中的磷酸结合,在结合过程中也产生H[SB]+[/SB](3Ca[SB]2+[/SB]+2HPO[XB]4[/XB][SB]2-[/SB]→Ca[XB]3[/XB](PO[XB]4[/XB])[XB]2[/XB]+2H[SB]+[/SB])。酸中毒可直接或间接破坏膜系统的功能和结构。
(3)氧自由基的产生休克时氧自由基产生增多主要是由于①氧代谢途径改变:即休克时由于细胞的缺氧和/或内毒素对线粒体呼吸功能的直接抑制,细胞色素氧化酶系统功能失调,以致进入细胞内的氧经单电子还原而形成的氧自由基增多而经4价还原而形成的水减少;②休克时产生大量乳酸、NADH及由ATP分解产生的次黄嘌吟等物质都可提供电子,使氧发生不全性还原而变成氧自由基。另外,休克时因蛋白水解酶活性增高,可催化黄嘌吟脱氢 酶变为黄嘌呤氧化酶,从而使次黄嘌吟变成黄嘌呤和氧自由基。③感染性炎症,活化补体等可激活中性粒细胞和巨噬细胞,使之释放出氧自由基。
氧自由基可通过膜脂质过氧化反应而破坏生物膜(参阅《缺血与再灌注损伤》)。
此外,溶酶体酶、内毒素等也可破坏细胞膜的功能与结构。
由于细胞膜的完整性在维持细胞的生命活动中起着重要作用。故当膜完整性破坏时,即意味着细胞不可逆性损伤的开始。
2.线粒体损害休克时线粒体最早出现的损害是其呼吸功能和ATP合成受抑制,线粒体ATP酶活性降低。此后发生超微结构的改变,如基质颗粒减少或消失;继之,基质电子密度增加、嵴内腔扩张,随后,嵴明显肿胀,终至破坏。
关于休克时线粒体损害的原因尚不完全清楚。缺氧可减少线粒体合成ATP,但除非在严重缺氧和伴有缺血时,并不引起线粒体膜的明显损害。目前认为,线粒体损害可能与下列因素有关:①内毒素等毒性物质及酸中毒对线粒体各种呼吸酶的直接抑制;②缺血导致线粒体合成ATP的辅助因子(如NAD、CoA和腺苷等)不足和细胞内环境(pH、离子)的改变。③前述的氧自由基对线粒体膜磷脂的过氧化作用等。
线粒体是维持细胞生命活动的“能源供应站”。线粒体损害时,由于氧化磷酸化障碍,产能减少乃至终止,故必然导致细胞损害和死亡。
3.溶酶体破裂溶酶体含有多种水解酶,如组织蛋白酶、多肽酶、磷酸酶等,但在未释放之前都处于无活性状态。一旦释放出来后,它们即转为活性状态而可溶解和消化细胞内、外的各种大分子物质,尤其是蛋白类物质。已证明,休克早期,肝、脾、肠等细胞即出现溶酶体肿大,颗粒丧失和酶释放增加;内毒素休克动物血液和淋巴中水解酶浓度增高,且与休克严重程度呈正相关。给动物注射溶酶体或溶酶体酶,可产生类似休克的各种病理生理改变。
休克时导致溶酶体破裂的主要原因是:①组织的缺血、缺氧、酸中毒以及内毒素对溶酶体膜的直接破坏;②氧自由基对溶酶体膜磷脂的过氧化作用;③血浆补体被激活产生C5a,后者可剌激中性粒细胞释放溶酶体酶。释放的溶酶体酶又可通过多种途径参与休克的发生、发展和细胞的损害,例如:a.释放的组织蛋白酶使蛋白质水解,这不但可以破坏蛋白酶的活性,甚至还可使细胞自溶坏死,而且所产生的多肽类活性物质,还能加重微循环障碍;b.破坏生物膜的完性;c.直接损害血管内皮和血管平滑肌细胞,从而导致血液外渗、出血和血小板的粘附、聚集以及DIC形成;d.激活补体系统产生C5a,后者再进一步促使溶酶体酶的释放。现已证明,休克时使用溶酶体膜稳定药可防止或减轻溶酶体膜的破裂。
总之,休克时生物的损害被认为是细胞发生损害的开始,而细胞的损害又是各脏器功能衰竭的共同机制。
四、器官功能的改变
休克时各器官功能都可发生改变,其中主要是中枢神经系统、心、肾、肺、胃肠及肝脏等重要器官的功能障碍。
(一)中枢神经系统功能的改变
休克早期,如果能通过代偿性调节维持脑的血液供给,除因应激反应而有兴奋性升高外,一般没有明显的脑的功能障碍。休克进一步发展,心输出量减少和血压降低,不能维持脑的血液供给,则发生缺氧。严重的缺氧和酸中毒还能使脑的微循环血管内皮细胞和小血管周围的神经胶质细胞肿胀,致脑微循环狭窄或阻塞,动脉血灌流更加减少。在微循环凝血期,脑循环内可以有血栓形成和出血。大脑皮层对缺氧极为敏感,当缺氧逐渐加重,将由兴奋转为抑制(表情淡漠),甚至发生惊厥和昏迷。皮层下中枢因严重缺氧也可发生抑制,呼吸中枢和心血管运动中枢兴奋性降低(详见《缺氧》)。
(二)心脏功能的改变
除心源性休克伴有原发性心功能障碍外,其它各类型休克也都可引起心功能的改变。一般而言,休克的早期可出现心的代偿性加强,此后心脏的活动即逐渐被抑制,甚至可出现心力衰竭,其主要机制是:
1.冠脉血流量减少和心肌耗氧量增加 由于休克时血压降低以及心率加快所引起的心室舒张期缩短,可使冠脉灌流量减少和心肌供血不足;同进因交感-儿茶酚胺系统兴奋使心率加快、心缩加强,导致心肌耗氧量增加,因而更加重了心脏缺氧。结果心肌因能量不足和酸中毒而使舒缩功能发生障碍,并从而引起心力衰竭,对于原来就有冠状动脉供血不良者,尤其容易出现心力衰竭。
2.酸中毒和高钾血症 酸中毒可通过多种机制影响心脏舒缩功能:①抑制肌膜的Ca[SB]2+[/SB]内流;②H[SB]+[/SB]和Ca[SB]2+[/SB]竞争结合肌钙蛋白;③抑制肌浆网对Ca[SB]2+[/SB]的摄取和释放;④抑制肌球蛋白ATP酶的活性。此外,酸中毒还可通过抑制心肌细胞能量代谢酶的活性、促使生物膜的破坏以及诱发心律失常等多种途径来抑制心肌的舒缩功能,并从而促使心力衷竭的发生。
休克时,组织细胞的破坏可释出大量K[SB]+[/SB],肾功能的障碍又使K[SB]+[/SB]的排出减少,因而总是伴有高钾血症。高血钾可抑制动作电位复极化2期中Ca[SB]2+[/SB]的内流,从而使心肌兴奋-收缩偶联减弱。
此外,心肌内DIC形成,内毒素对心肌的直接作用等等。都可以促使心肌力衰竭的发生。一旦发生了心力衰竭,将迅速促使休克进一步恶化,并给输液扩容造成一定困难。
3.心肌抑制因子的作用如前文所述,休克时的缺血、缺氧等可使胰腺产生心肌抑制因子(MDF),MDF能使心肌收缩力减弱,从而有助于心力衰竭的发生。
(三)肾功能的改变
肾功能的改变在休克早期就可发生,这时发生的是功能性的急性肾功能衰竭,因为它还不伴有肾小管的坏死。其主要临床表现为少尿(<400ml/d)或无尿(<100ml/d),其发生的主要机制如下:
1.肾小球滤过率减少在休克早期,有效循环血量的减少不仅能直接使肾血流量不足,而且还可通过肾素-血管紧张素系统和交感-儿茶酚胺系统的激活而使肾血管收缩,因而使肾血流量更加减少,结果是肾小球滤过压降低,肾小球滤过率减少。
2.肾小管对钠、水重吸收加强在休克早期,肾小管上皮细胞虽然已经发生缺血,但是因为持续时间不久,故这些细胞仍能保持其正常的重吸收功能,加之此时醛固酮和抗利尿激素分泌增多,所以肾小管对钠水的重吸收加强。肾小球滤过率的减少和肾小管重吸收的增强就可导致少尿或无尿。但此时肾功能的变化是可逆的。一旦休克逆转,血压恢复,肾血流量和肾功能即可恢复正常,尿量也将随之而恢复正常。故尿量变化是临床判断休克预后和疗效的重要指标。
当休克持续时间较长时,可引起急性肾小管坏死,发生器质性的肾功能衰竭。此时即使肾血流量随着休克的好转而恢复,患者的尿量也难以在短期内恢复正常。肾功能的这些改变,将导致严重的内环境紊乱,包括高钾血症、氮质血症和酸中毒等。这样就会使休克进一步恶化,故许多休克患者,尤其是老年患者常死于急性肾功能衰竭。
(四)肺功能的改变
随着休克的发展,肺功能也发生不同程度的改变:在休克早期,由于呼吸中枢兴奋,故呼吸加快加深,通气过度,从而甚至可以导致低碳酸血症和呼吸性碱中毒;继之,由于交感-儿茶酚胺系统兴奋和其他血管活性物质的作用,可使肺血管阻力升高;如果肺低灌流状态持续较久,则可引起肺淤血、水肿、出血、局限性肺不张、微循环血栓形成和栓塞以及肺泡内透明膜形成等重要病理改变,此即所谓休克肺(shock lung)的病理学基础(参阅第十三章)。
上述休克肺的病理变化,有的影响肺的通气功能。有的妨碍气体弥散,有的改变肺泡通气量/血流量的比例,造成死腔样通气和/或功能性分流,从而可以导致呼吸衰竭甚至死亡。休克肺是休克死亡的重要原因之一,约有1/3的休克患者死于休克肺。
(五)肝和胃肠功能的改变
1.肝功能的改变休克时常有肝功障碍,其主要原因有:①低血压和有效循环血量减少可使肝动脉血液灌流量减少,从而引起肝细胞缺血缺氧,严重者可导致肝小叶中央部分肝细胞坏死;②休克时由于腹腔内脏的血管收缩,致使门脉血流量急剧减少。肝约有一半以上血液来自门脉,故门脉血流量减少,也将加重肝细胞的缺血性损害;③肝内微循环障碍和DIC形成,更可引起肝细胞缺血缺氧;④在肠道产生的毒性物质经门脉进入肝,加之肝本身毒性代谢产物的蓄积对肝细胞都有直接损害作用。
肝功能障碍又可通过下列机制加重休克:①肝代谢障碍:肝对糖和乳酸的利用障碍,一方面可促使乳酸蓄积并从而引起酸中毒;另方面又不能为各重要脏器提供充足的葡萄糖。蛋白质和凝血因子合成障碍,可引起低蛋白血症和出血。②肝的生物转化作用(解毒功能)减弱:可增加休克时感染与中毒的危险。
2.胃肠功能的改变 休克早期就有胃肠功能的改变。开始时是因微小管痉挛而发生缺血,继而可转变为淤血,肠壁因而发生水肿甚至坏死。此外,胃肠的缺血缺氧,还可使消化液分泌抑制,胃肠运动减弱。有时可由于胃肠肽和粘蛋白对胃肠粘膜的保护作用减弱,而使胃肠粘膜糜烂或形成应激性溃疡。
由于胃肠上述改变,可通过下列机制促使休克恶化:①肠道粘膜屏障功能减弱或破坏,致使肠道细菌毒素被吸收入血,加之肝的生物转化作用减弱,故易引起机体中毒和感染。②胃微循环淤血,血管内液体外渗,加之胃肠粘膜糜烂坏死和DIC的形成都可导致胃肠道出血,从而使血容量进一步减少。③胃肠道缺血、缺氧,可剌激肥大细胞释放组胺等血管活性物质,因而微循环障碍进一步加剧。
(六)多器官功能衰竭
多器官功能衰竭(mrltiple organ failure MOF)是指心、脑、肺、肾、肝、胃肠、胰腺及血液等器官中,在24小时内有两个或两个以上的器官相继或同时发生功能衰竭。MOF又称多系统功能衰竭或综合器官衰竭。休克的晚期常出现MOF。MOF是致死的重要原因,而且衰竭的器官越多,病死率也就越高。如有三个器官发生功能衰竭时,病死率可高达80%以上。
MOF在临床上有两种表现形式,一是创伤和休克直接引起的速发型,又称单相型,发生迅速,发病后很快出现肝、肾和呼吸功能障碍,在短期内或则死亡,或则恢复;二是创伤、休克后继发感染所致的迟发型,又称双相型。此型患者往往有一个相对稳定的间歇期,多在败血症发生后才相继出现多器官功能衰竭。引起MOF的主要原因是:①重症感染:约有70~80%的MOF是在重症感染的基础上发生的;②休克时组织较长时间的低灌流和交感神经的高反应;③非感染性的严重病变如急性胰腺炎、广泛性组织损伤等。尤其是当机体的免疫功能和单核吞噬细胞系统功能减弱时,或者是治疗不当或延误时,如未及时纠正组织低灌流和酸碱平衡紊乱、过多过快输液、大量输血或过量应用镇静剂、麻醉剂等情况下,更易引起MOF。
MOF的发病机制尚不很清楚,现认为MOF的发生是多因素参与作用的结果,其中休克时组织低灌流所致的组织缺血缺氧、代谢障碍和酸中毒都起着重要作用;在感染或感染中毒性休克时,细菌内毒素在MOF的发生机制中被认为是起着关键的作用。这是因为内毒素不但能直接或间接(如激活补体和凝血系统)损害各器官的功能,而且还可通过激活补体而使中性粒细胞聚集和激活,从而使中粒细胞①释放各种水解酶,包括酸性和碱性蛋白酶,这些酶不但能破坏结构蛋白(如弹性蛋白酶破坏弹性纤维,胶原酶破坏胶原纤维等),而且还可分解血浆蛋白,激活凝血系统并从而导致弥散性血管内凝血;②产生并释放活性氧和脂类代谢产物(如白三烯等),这些物质又可破坏生物膜和/或增高血管通透性,加重微循环障碍。在一般情况下,中性粒细胞向感染和损伤处趋化和聚集,是炎性的正常反应,但在休克或严重感染时,由于机体免疫功能降低等原因,对这种炎性反应失去控制,从而使中性粒细胞释放的上述各种毒性物质得以广泛地破坏各器官细胞的结构和功能。故有人认为MOF是一种非特异性失控的全身性炎性反应。此外,儿茶酚胺-腺苷酸环化酶-cAMP系统异常也可能起着重要的作用,在休克时一方面因为细胞的缺血缺氧,使膜功能异常,腺苷酸环化酶系统的受体受损,对儿茶酚胺的反应减弱;另方面由于组织ATP含量降低,缺乏产生cAMP的底物,结果使细胞内cAMP水平下降,进而影响细胞内的许多代谢过程和功能。