第一节 免疫学研究中实验动物的选择与应用
一、免疫学研究中实验动物的作用
免疫学是近年来医学科学中发展较快的一门学科。随着分子生物学、免疫化学等的发展和免疫学新技术的应用,免疫学得到了飞跃发展,不仅改变了许多旧的传统观察,而且由于免疫学的知识已渗透到医学科学的各个领域,形成了多种新兴学科,给许多疾病的诊断、预防及治疗开辟了广阔的前景。现在在医学理论与医学实践中,涉及有关免疫学的问题越来越多,受到人们广泛的重视。
免疫学的发展与实验动物科学兴起有密切关系。免疫学的研究,包括从预防感染到区别机体自身或非自身的基本生物现象有研究,一般多选用实验动物作对象,而且免疫学上的大量知识是通过动物实验获得的。特别是各种近交系和突变系动物、无菌动物、悉生动物及无特定病原体动物的培育,为免疫学研究提供了重要手段,大大促进了免疫学的发展。
为了研究免疫反应的原理以及发病机理的本质,单从人体内进行研究是非常困难的,我们观察病人,无法了解他的遗传背景,他过去和现在所处的环境,检查前的免疫状况以及今后发展的过程。以老年的免疫反应来讲,虽然许多人对老年时的免疫状态作了许多工作,但由于个体差异非常之大,单纯用平均数很难说明问题,因为只能代表一个趋势,而不是每个个体的真正反应。在这一群人中间,过去的基础不相同,因而不能同过去比较,就是用平均数字也不能肯定当时的水平,无法确定,是结果还是老年的原因。各种免疫病也是如此。因此,要真正了解免疫的本质,就不能单在病人身上或取血做体外实验,应主要在动物模型上进行研究,才能克服上述困难,取得更多真正能解决根本问题的突变性研究成果。
二、影响实验动物免疫反应的因素
免疫反应是随着动物种的进化而复杂化、精密化和完善化的,不同品种和品系动物的免疫反应有很大差异,因此在免疫学研究中选择动物时要特别注意动物遗传因素的影响,还要注意动物的年龄因素、感染因素、营养因素、药物因素、母源因素、应激因素、免疫抑制剂等因素对动物免疫反应的影响。
(一)动物的遗传因素
根据研究表明小鼠、豚鼠、家兔等动物对特异性抗原的免疫反应受遗传控制。动物体内免疫反应的基因决定着动物对各种疾病的易感性,决定着自身免疫病和体液免疫反应。这种免疫反应的基因紧密连接在这些动物体内主要组织兼容系统上。如带等位基因H-2b的小鼠(如C57BL,C57L,129/J)比带有等位基因H-2K的小鼠(如C58,AKR,C3H)的抵抗力强,后者对小鼠白血病病毒和肿瘤病毒十分易感,又如SWR/J(H-2q)小鼠对淋巴细胞性脉络丛脑膜炎病毒(LCM)非常敏感,而C3H/J(H-2k)小鼠对该病毒有强大的抵抗力。这些例子说明由于遗传因素的影响,不同品系动物的免疫反应是有明显差异的。此外,不同种类动物的免疫反映也有差异,如研究第Ⅳ型变态反应(Arthus反应),家兔是一种好的实验动物,而豚鼠和大鼠不能采用。豚鼠通常产生少量的IgM。
脊椎动物免疫系统的发生与种系化密切有关(见图10-1、2和表10-1):原始脊椎动物的淋巴器官发育还不完善,如圆口类动物沿其消化道有散在的淋巴结和淋巴细胞,并出现了胸腺,随着进化有了原始的肾脏,在鱼类还出现了肝脏。这些器官和组织开始时也多分布在消化道附近,这是由于原始脊椎动物(圆口类)及鱼类摄食时吸进大量水,并通过鳃孔将水排出,因此咽头部最先遭受病原微生物的侵击,为此在消化道附近产生了相应的御体系。而到了高等脊椎动物,由于种系的进化,这些器官的分布就多样化了。但从高等动物胸腺个体发生来看,它也是从第三、第四咽囊腹侧上皮演化发育而来,说明这跟种系发生有关。
虽然圆口类动物就能产生IgM,但无控制B淋巴细胞分化发育的中枢器官,至两栖类开始出现较原始的类囊器官。而爬行类动物的类囊,位于消化道的末端。鸟类以外其他动物有类体液免疫完善的免疫系统。
因此,在免疫学研究中进行实验动物选择时,要特别注意遗传因素对免疫反应的影响,各种实验动物具有不同的免疫反应和免疫特点。下面再举个典型例子。实验动物补体系统各成分的缺陷因实验动物的种类不同而有明显差异。
补体缺陷(Complemenr Deficiencies)
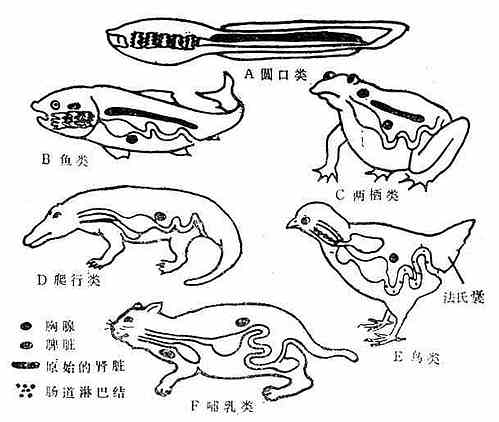
图10-1 脊椎动物的免疫反应
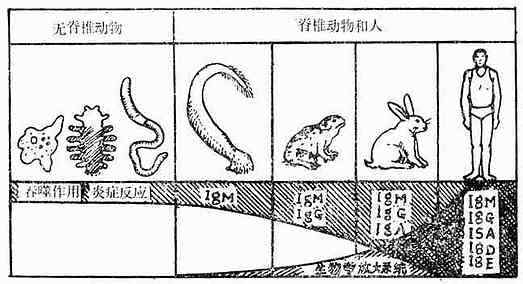
图10-2 免疫应答反应的种系发育主要代表
C1 鸡
C3 狗(Brittary Spaniel)
C4 金黄地鼠
C5 小鼠(K/HeN.AKR/N、B10、DZ/DsnN)
C6 兔、地鼠
当C1g缺乏时可出现严重的联合性免疫缺乏病,反复发生威胁生命的感染;C1r缺乏时发生坏疽性红斑,反复的细菌感染,狼疮样综合症;C1s缺乏时,出现红斑狼疮,进行性肾小球肾炎,关节炎;C4缺乏时可发生狼疮,关节炎、类过敏性紫癜;C2缺乏时发生狼疮,致死性皮肌炎,类过敏性紫癜,狼疮样综症,进行性肾小球肾炎,反复感染;C3缺乏时对感染的易感性升高;C5缺乏时可发生狼疮,腹泻及消耗病:C6缺乏时,可发生反复G(-)感染,淋菌性多关节炎,反复发生脑膜炎。
| 动 物 | 细 胞 | 淋 巴 器 官 | 免 疫 反 应 | ||||||||||
| 分类 | 动物
名称 | 淋巴细胞 | 浆细胞 | 胸腺 | 脾脏 | 淋巴结 | 法氏囊 | 细胞免疫 | 体 液 免 疫 | ||||
| IgM | IgG | IgA | IgE | IgD | |||||||||
| 圆口类 | 鳗 | + | - | + | - | - | - | + | + | - | - | - | - |
| 鱼类 | 油鲛 | + | - | + | + | - | - | + | + | - | - | - | - |
| 鲤鱼 | + | + | + | + | - | - | + | + | - | - | - | - | |
| 两栖类 | 蝾螈 | + | + | + | + | - | + | + | - | - | - | - | |
| 青蛙 | + | + | + | + | (+) | +* | + | + | + | - | - | ||
| 爬行类 | 龟蛇 | + | + | + | + | (+) | ? | + | + | + | - | - | - |
| 鸟类 | 鸡 | + | + | + | + | + | + | + | + | + | +? | - | - |
| 哺乳类 | 鼠、人 | + | + | + | + | + | +* | + | + | + | + | + | + |
(+)可能存在,+*有功能相似的类囊器官。
图10-3 各种动物胸导管淋巴细胞数目的变化(细胞数/公斤体重)
(二)动物的年龄因素
免疫学研究中选择适宜年龄的实验动物是非常重要的,年龄影响着实验动物的免疫机能。如幼龄动物的免疫系统机能发育不完善或机能很弱。一般来说,动物的免疫机能在青年期达到高峰,以后随着年龄增大逐渐减弱,主要表现有血清中免疫球蛋白含量低,细胞免疫功能下降,恶性肿瘤和自身免疫性疾病的发病率增高等。据研究,小鼠、大鼠和豚鼠随年龄增加免疫反应的活性也减弱,老龄鼠产生IgG和IgM的能力仅为年青成年鼠的10%左右,细胞免疫同样也减弱。因此,老龄鼠对诱发肿瘤极敏感。
实验证明给胚胎期或新生期的动物注射异基因型细胞时很容易造成对该细胞的免疫无反应性(Immunolgical Unresponsivenss),而对成年动物注射异基因型细胞时通常可引起免疫反应。据认为这种免疫耐受性与免疫系统(包括中枢淋巴样器官即骨髓、胸腺和外周淋巴样器官即脾脏、淋巴结、肠管相关淋巴样组织以及循环的淋巴样细胞)的发育未臻成熟有关。
Heidrick和Makindan氏认为老龄鼠细胞免疫中免疫细胞的缺乏,可能是由于T细胞和B细胞没有能力增殖的缘故。通常以为动物越是趋于老年,免疫反应的自隐机制被破坏的机会就越多,因而发自身免疫疫病的机会就越多。这种自稳对动物的正常生命活动乃至生存具有重要意义。在正常情况下,健康机体对自己的细胞、非细胞成分如蛋白质、多糖等是不发生“明显的”免疫反应的(自身免疫耐受状态,Selftolerance),而对非自己的抗原则发生免疫反应,也就是说机体通过免疫系统能识别“自己”和“非自己”。这种自我识别能力称之为自稳机制。
(三)动物的母源因素
各种哺乳类动物的胎儿和出生仔畜免疫的获得不同。在初乳中主要为IgA,初生动物血清中的母源抗体大部分为IgG,对仔畜预防病毒和细菌感染起着一定的保护性免疫作用。大部分IgA对粘膜表面起着局部保护作用,而IgG起着总的保护作用。但是母源抗体还有有害作用,它能诱发新生动物溶血病(如驹)和抑制出生动物的主动免疫。
雌性动物将母源抗体转移给胎儿或仔畜的途径和特异性不同,这与动物胎盘的结构和类型有关(见下页表10-2),一般来说,有三种主要的转移途径,随动物种类不同,有的经绒毛膜尿囊胎盘转移;有的经卵黄囊上皮和卵黄循环转移;还有是初乳经肠道吸收。前两者被认为是胎儿期获得抗体的途径,后者为出生后转移抗体的途径。
免疫球蛋白的转送是有选择性的,有些种类的抗体易转移,同种(系)抗体转移比异种抗体快。胎盘对各种母源抗体也有选择性,如:灵长目中Igg 易通过胎盘屏障,IgM、IgA和IgE则不能。家兔的IgG和IgM易通过胎盘到达胎儿。关于母源抗体选择性的转移机制还不清楚,一般认为,这种选择性的转移是由于组织能选择性地识别抗体的F。端造成的,母源抗体转移中具有选择作用的组织见表10-3。动物常乳和初乳中检出抗体的类型见表10-4。
表10-2 动物的胎盘屏障和母源抗体的转移
| 动 物 | 胎盘类型 | 结构(层数) | 转移途径 | 选择性 | 抗体转移 | 持续时间 | |
| 出生前 | 出生后 | ||||||
| 小鼠 | 血性绒毛膜 | 4 | 肠(主要)卵黄囊 | + | +(卵黄囊) | 艹 | 出生到17天 |
| 大鼠 | 血性绒毛膜 | 4 | 肠(主要)、卵黄囊 | + | +(卵黄囊) | 廿 | 出生到20天 |
| 豚鼠 | 血性绒毛膜 | 2 | 卵黄囊(主要)、肠 | ? | 卅 | + | 整个妊娠期间和出生后2天内 |
| 家兔 | 血性绒毛膜 | 2 | 卵黄囊 | + | 卅 | - | 从妊娠开始15天起 |
| 灵长类 | 血性绒毛膜 | 2 | 胎盘 | + | 卅 | 卅 | 妊娠开始3个月 |
| 猬 | 肠 | + | - | 艹 | 出生到35天 | ||
| 狗 | 内皮绒毛膜 | 4 | 肠 | ? | - | 卅 | 出生到8天 |
| 猫 | 肠 | ? | - | 卅 | 不定 | ||
| 水貂 | 肠 | ? | - | 卅 | 出生到8天 | ||
| 雪貂 | 肠 | ? | - | 卅 | 出生到34天 | ||
| 猪 | 上皮绒毛膜 | 6 | 肠 | - | - | 卅 | 出生后36小时 |
| 马 | 上皮绒毛膜 | 6 | 肠 | - | - | 卅 | 出生后36小时 |
| 牛 | 结缔组织绒毛膜或上皮绒毛膜 | 6 | 肠 | - | - | 卅 | 出生后36小时 |
| 绵羊 | 结缔组织绒毛膜或上皮绒毛膜 | 6 | 肠 | - | - | 卅 | 出生后36小时 |
表10-3 母源抗体转移中呈现选择作用的组织
| 动 物 种 类 | 组 织 |
| 灵 长 目 | 胎 盘 |
| 大鼠与小鼠 | 肠 |
| 家 兔 | 卵 黄 囊 |
| 偶 蹄 兽 | 乳 腺 |
| 鸟 | 卵黄囊膜 |
表10-4 动物常乳和初期中免疫球蛋白类型
| 动 物 | 常乳和初乳中免疫球蛋白类型 | |
| 家 兔 | 初乳 | IgA |
| 灵长目 | 初乳 | IgA |
| 大 鼠 | 初乳 常乳 | IgA(多量)IgGa,IgGb,r1,IgM(很少) |
| IgG(多量) | ||
| 小 鼠 | 初乳 | IgA,快和慢的IgG |
| 狗 | 初乳 常乳 | IgG(多量),IgA,IgM |
| IgA(多量),IgG,IgM | ||
| 猪 | 初乳 常乳 | IgG(多量),IgA,IgM |
| IgA(多量) | ||
| 牛 | 初乳 | 快IgG(r1)IgM(β2) |
| 绵 羊 | 初乳 | 快IgG(r1),IgA |
(四)动物的感染因素
病毒、细菌、真菌或寄生虫的感染都能多方面地改变动物机体的生理功能,这种改变无疑将影响到动物的免疫系统。动物感染可引起继发性营养不良,从而影响到免疫反应。一般实验动物常发生病毒的隐性感染,在以下几方面影响免疫功能:抗体产生的方式,免疫球蛋白的数量,免疫耐受性的产生,植皮排斥,植皮对宿主的反映,迟发型变态反应,淋巴细胞转化和吞噬作用等。
动物病毒感染引起的免疫抑制表现:干扰正常免疫系统的功能;改变抗原的吸收和处理;破坏抗体形成细胞和浆细胞(如白血病病毒)。病毒感染后也可引起免疫增强的表现,如乳酸脱氨酶病毒LDH,委内瑞拉马脑脊髓炎病毒VEE等病毒,能增加产生抗体细胞数量:感染病毒的细胞能产生微量核苷酸,起着佐剂的作用;病毒还能促进免疫细胞对抗原的处理。
动物感染呼吸道病毒、腺病毒、流行性脊髓灰质炎病毒、仙台病毒、痘苗病毒、单纯疮疹病毒、新城疫病毒等病毒时,能够抑制淋巴细胞的转化。肝炎、肢骨发育不全(Ectromelia)、淋巴细胞性脉络丛脑膜炎(LCM)、乳酸脱氨酶病毒(LDH)能抑制网内皮系统的机能。而Friend白血病、Moloney白血病和委内瑞拉马脑脊髓炎(VEE)能增强吞噬细胞的吞噬作用。
三、实验动物免疫反应的特点
(一)灵长类(除人外)
灵长类动物主要有四种免疫球蛋白,即IgG、IgM、IgA、和IgE。新界猴(除一种卷尾猴外)没有发现IgA。已证明在猕猴、狒狒和黑猩猩中有抗寄生虫性抗原的IgE抗体,但在新界猴中仍无此种抗体。高等灵长类与人的免疫球蛋白有较强的交叉反应,但长臂猴例外。
灵长类动物具有血性绒毛膜胎盘,只允许IgG通过,IgM、IgA、IgD和IgE是不能通过的。新生的猴子不能从初乳中吸收抗体。妊娠第九个月猕猴的胎儿和成年猕猴在抗原初次刺激后6天产生IgM,妊娠第58天的胎儿对同种植皮产生排斥,而羊胎儿的这种反应发生在80天。
80%出生的狨猴是胎盘血管吻合的双胎。用性别染色体分析能证明不同性别的双胎中血液存在交换。在血液交换的狨猴双胎中,异性共生的双胎已证明有免疫耐受现象。因此,它们之间互相接受植皮。狨猴对接受不同亚种的植皮有免疫反应,亚种内植皮要比亚种间植皮存活的时间约长一倍。
灵长类动物在研究人的反应素(IgE)型超敏反应中起着重要作用。反应素型抗体(又叫皮肤过敏性抗体)特点之一,是能固定在同源或密切相关种的皮肤及其它组织上(如:肺、结肠)。由于猕猴同人在种的发生上有近缘关系,所以它们能用过敏人的血清引起P-K反应(Prausniz-Kusrner)。在灵长类中,狒狒、猕猴、狨猴、卷尾猴、狐猴是人类过敏性患者反应素抗体引起的萎缩性多软骨炎(PCA)的最好接受者。一些学者证明,灵长类动物是人呼吸道变态反应病的动物模型。
(二)狗
狗的免疫球蛋白有IgG、IgM、IgA、IgG[XB]1[/XB]和IgG[XB]2[/XB]。在狗花粉病和各种蠕虫感染中发现有IgE。Patterson氏等指出成年狗对各种蛋白抗原只产生少量的循环抗体。胎儿和新生狗也有类似情况。
Lewis等认为新生狗和成年狗对颗粒性抗原(绵羊红细胞)均能产生较好抗体,但新生狗初次免疫反应所产生的抗体几乎是IgM类,成年狗则产生的抗体是IgM和IgG,这两种免疫球蛋白的数量与初生狗的IgM几乎相等。新生狗在再次反应中有合成IgG和IgM。
Gerber氏等报告了小猎兔犬(Beagle Dog)循环的T淋巴细胞对PHA的反应,6-12周龄比0-4周龄显著为多。对PHA发生反应的高峰在6周到6个月龄之间,以后随年龄而下降。小猎兔犬在出生时的胸腺大约为100mg,到12周龄增加到300mg以上。白细胞总数随年龄增加而逐渐减少。
狗除用作一般移植研究外,越来越多地作为免疫病研究的动物模型。除人之外,对气溶胶出现变态反应的动物,狗大概是仅有的一种。因此,人的变态反应和气喘的研究,狗是适宜的动物模型。人花粉病的临床表现为结膜炎、鼻炎和皮炎,狗季节性花粉病多数只有皮炎,无眼和呼吸道症状。人的这种变态反应是由IgE引起的,狗由豚草花粉(Raweed Pillens)致敏后,血液和皮肤中也有IgE抗体。
(三)兔
兔常被用于实验室制备抗体,多用新西兰色。由于所用兔的品系、品种、种和个体的不同,对某种抗原产生抗体的能力也经常不同。有些品系的兔,至少有20%产生的抗体效价或无效价,为了得到高效价的血清,10只兔作为一组进行免疫要必要的。
兔肠道淋巴组织由派伊尔氏淋巴集结,圆囊(在回盲肠连接处的集合淋巴小结)和阑尾构成。Waksman氏等叙述了淋巴细胞组织中形态和机能不同的三个部分:圆体(Dome)含有原始和成熟的B淋巴细胞;冠(Corona)含有小淋巴细胞胸腺依赖区含有后毛细菌小静脉、滤泡。他认为圆体可能相当于囊淋巴器官,能产生B淋巴细胞。滤泡是一种非特异性的B细胞大量增殖(扩大)的器官。集合淋巴小结、圆囊和阑尾都含有T细胞,T细胞经过后毛细胞管静脉到达阑尾。集合淋巴结和阑尾内还同时存在着B细胞,但没有抗体形成细胞。这种界线分明的T、B淋巴细胞系统及B细胞迅速地增殖和迁移(肠道淋巴器官中,B细胞参加免疫反应,但不长久停留)可能使肠道局部缺乏免疫反应。集合淋巴小结是产生IgA细胞的重要来源。
兔的IgA大量存在于肠粘膜和初乳中,这种分泌型抗体的合成是在肠、乳房和支气管腺体间质的浆细胞以及脾和淋巴结中。
兔的反应素抗体相当于人的IgE。兔的IgM能增强反应素的形成,而IgG能抑制反应素抗体的生成。
兔被用来作过敏反应的研究。IgG和IgE引起的过敏反应,临床症状相似,机理都是抗原-抗体结合和血小板-白细胞凝集形成沉淀物,释放药理活性物质(组织胺和5-羟色胺)进入肺循环,在右心的出道中产生一种机械和药理的联合作用,导致循环性虚脱。IgG诱发血小板或嗜碱性的细胞释放影响血管的胺要依赖补体的作用,而IgE诱发释放的胺不依赖补体。
(四)豚鼠
豚鼠中已确定的免疫球蛋白有:IgG(IgG[XB]1[/XB]、IgG[XB]2[/XB])、IgA和IgE。IgG[XB]1[/XB]是变态反应的媒介,IgG[XB]2[/XB]与小鼠的IgG[XB]1[/XB]和IgG[XB]2[/XB]相似,在抗原一抗体作用中起结合补体的作用。
豚鼠除作为补体的来源外,已广泛用于免疫的发生和迟发型变态反应的研究。近新繁殖的豚鼠2系和13系常被用于免疫学研究。这两个品系对特异性抗原产生的免疫反应有显著不同。例如,当给豚鼠2系和13系注射含有相同抗原的福氏(Freund)完全佐剂时,豚鼠2系(和一些Hartley系豚鼠)表现明显的迟发型变态反应,对DNP-PLL(二硝基苯-多-L-溶解素)产生高浓度的抗体,而豚鼠13系不出现免疫学反应。另一方面,豚鼠13系和Hartley系豚鼠对联胺嗪(Hydralazine)都能产生抗体和迟发型变态反应。豚鼠2系仅呈现弱反应或无反应。
豚鼠的皮肤已被用已结核菌素的皮内试验和接触过敏物质的迟发型变态反应的研究。豚鼠和人的结核菌素反应差别是有无细胞浸润,另外豚鼠的迟发型变态反应在24-48小时达到高峰,人在48-96小时达到高峰;人和豚鼠接触敏感的化学物质引起的变态反应,细胞反应非常相似,而对皮内接种抗原的反应却有明显的不同,豚鼠比人有更多的白细胞和巨噬细胞对抗原起反应。
当进行免疫学研究选择豚鼠时,应特别注意机体本身的因素,如:年龄、体重、饮食和遗传因素。Baer氏等认为2-3月龄或体重为350~400克的豚鼠作迟发型变态反应最合适。豚鼠13系对结核菌素型变态反应比豚鼠2系敏感。相反,豚鼠2系对接触性过敏反应比豚鼠13系敏感。Hartley系豚鼠对结核菌素型变态反应和接触反应皆敏感。这些现象说明抗体发生迟发型变态反应的能力同样也处于基因的控制之下。
最近,一些学者以豚鼠作为研究过敏或速发型过敏反应的实验模型。在全身变态反应中,肺是休克器官,肥大细胞是靶细胞,组织胺是主要的药理价质。在豚鼠在两种类型的变态反应抗体,即IgG1和IgG2。
Recently Arko报导,在进行淋病研究中使用的实验动物,豚鼠是最令人满意的免疫学模型。豚鼠象人一样具有延长和限制迟发型真皮变态反应显现的能力,这种现象常作为肿瘤免疫的指标之一。
(五)仓鼠
Coe氏等研究了Syrian仓鼠的免疫反应,发现有电泳快的(IgG1)和慢的(IgG2)两种TS亚类免疫球蛋白。当以鸡蛋白盐水作为抗原,接种仓鼠,仓鼠产生IgG1;若将鸡蛋白与福氏佐剂一起接种仓鼠,则能产生IgG1和IgG2。仓鼠的IgG1能诱发PCA反应,不能产生全身过敏反应。Coe氏等认为这可能是由仓鼠在变态反应中缺乏必要的影响血管的胺的缘故。仓鼠的IgG2能固定补体,并在豚鼠中诱发PCA反应,IgG1不能固定补体。
(六)小鼠
小鼠的免疫球蛋白有IgM、IgA、IgE、IgG[XB]1[/XB]、IgG[XB]2[/XB]a和IgG[XB]2[/XB]b。近交系小鼠对不同抗原的免疫反应是在常染色体的遗传控制之下,这种常染色体上有支配免疫反应的基因(Ir),基因连接在主要组织相容位点(H-2)上。基因Ir好象同T细胞的机能有关系,与B细胞的关系不大。
小鼠虽然能产生迟发型变态反应,但很少见到典型的表皮反应,也不象其他动物那样有规律。小鼠能被诱发产生速发型变态反应,它的全身性过敏反应的特点是循环不畅,循环性虚脱,常在几小时甚至10到20分钟死亡。在体外过敏反应实验中,只有小鼠子宫能用来作Schultz-Bale反应。小鼠的IgG和IgE能使皮肤致敏,引起被动真皮过敏反应。诱发小鼠的Arthus反应比较困难,既使发生,与其他实验动物(如兔)相比也不那么激烈。小鼠不象大鼠和豚鼠那样,以福氏完全佐剂接种不小鼠的脊髓或脑内很难诱发实验性变态反应脑脊髓炎(EAE),用主要组织相容性综合体不能控制小鼠对实验性变态反应脑脊髓炎的感受性。
(七)大鼠
在大鼠中,连接在主要组织相容综合体(H)上的免疫反应基因(Ir)控制着对GT(L-谷氨酸和L-酪氨酸)和GA(L-谷氨酰胺和L-氨基丙酸)的免疫反应,豚鼠与其相似。
大鼠和豚鼠的免疫反应基因lr控制着体液抗体反应和细胞免疫。已经证明,大鼠对绵羊细胞(SRBC)和牛r球蛋白(BGG)的免疫反应有品系的差异。
大鼠有反应素抗体IgE,蠕虫感染常能诱发大量的IgE抗体,它们存在于血液循环之中。常规的免疫法只能使大鼠产生少量反应素,在体内存在的时间较短。有些品系大鼠,如Hooded Lister和Spragus-Dawley,能产生较多的IgE,再次注射抗原,IgE也随之上升。百日咳菌免疫大鼠主要产生IgE,如在此抗原中加入福氏完全佐剂,免疫大鼠则产生IgGa。
(八)猪
已经知道猪有三种免疫球蛋白,即IgG(IgG[XB]1[/XB]和IgG[XB]2[/XB])、IgM和IgA。猪初乳中的免疫球蛋白主要是IgG(其中IgG1为主),其次是IgA。泌乳2-3天后,乳中IgG和IgM迅速下降,但IgA的量仍保持相对稳定。
猪的IgA同人的IgA有交叉反应。IgA有单体和存在于分泌中的双体两种,它们分别为7s和10s。肠道固有层中包含着多量分泌IgA的浆细胞。
(九)反应动物(牛、绵羊、山羊)
反刍动物已经确定的免疫球蛋白有IgG(IgG[XB]1[/XB]和IgG[XB]2[/XB])、IgA、IgM和类似IgE。同其它动物一样,IgG是主要的免疫球蛋白,IgG[XB]1[/XB]能固定补体(IgG[XB]2[/XB]不能),并能选择性从血清到乳汁中去,所以初乳和常乳中IgG[XB]1[/XB]是主要的免疫球蛋白(75%),而IgA和IgM仅占初乳抗体的20%。这些初乳中的免疫球蛋白对于幼畜的存活是非常重要的,它们能协助胃肠道预防感染。
Silverstein氏等研究了羊免疫反应的个体发生,发现不同日龄的胎儿对抗原的反应有显著差别。41天的胎儿对细菌噬菌体×174能产生抗体,56天的胎儿对铁蛋白能产生抗体,80天的胎儿对血清蛋白能产生抗体,120天的抬儿对卯清蛋白能产生抗体。胎儿对卡价苗、伤寒沙门氏菌或白喉杆菌病毒没有反应,羊胎儿对抗原同样会产生典型的初次抗体反应。
四、近交系动物在免疫研究中的应用
(一)自身免疫性疾病
自身免疫性疾病主要的发病机理是免疫复合物造成的损害,如系统性红斑狼疮(SLE)、肾炎、类风湿性关节炎等,已知抗体起作用的疾病有重症肌无力、甲状腺机能亢进症等,此外还有一些目前不清楚的免疫病,即对其抗原和抗体的免疫反应性都不清楚,这些病研究起来比较困难。由于免疫复合物病有较好的动物模型,研究起来就方便多了。
1.研究感染与自身免疫病的关系。
纯系新西兰黑色小鼠(New Zealand、Black,NZB)在出生后4-6个月大多数发生自身免疫性溶血性贫血。免疫学检查表明,其红细胞表面有自身抗体,血清中有抗核机体,病鼠的淋巴结、胸腺、脾脏及胰腺内可以查到C型致瘤RNA病毒(此病毒还能引起白血病)。将含有此病毒的组织输给(NZB×BALB/c)F[XB]1[/XB]时,能引起抗核抗体的生成和肾小球肾炎。据推测,当宿主细胞受到病毒感染后,释放出DNA,刺激免疫系统生成抗DNA自身抗体,此抗体与游离的DNA结合,形成免疫复合物沉积在肾小球基底膜上,引起肾小球肾炎,因此认为本病是由于这种病毒感染所触及的。此外,还发现约有50%的病鼠有抗自身T淋巴细胞的抗体,且其出现频率随鼠龄增加而升高,这种自身抗体能与正常鼠及病鼠的T细胞发生反应,引起T细胞(包括抑制性T细胞)减少或功能衰退,结果,B淋巴细胞乃生成大量多种自身抗体。人的系统性红斑狼疮与NZB小鼠的这种自身免疫病十分相似,也能在淋巴样组织中查到C型RNA病毒颗粒,其T淋巴细胞也能释出病毒颗粒,也能查到抗自身T淋巴细胞的抗体(约占一半病例)而且病理变化也基本上与NZB小鼠的这种自身免疫病相同。
除了NZB小鼠的自身免疫性溶血性贫血以外,阿留申貂脑病(一种脑的退行性变性病)、小鼠的淋巴细胞性脉络丛脑膜炎、羊的瘙痒病(skrapie)、马的传染性贫血也都是由于慢性病毒感染引起的自身免疫病。人的Kuru病与羊的瘙痒病、阿留申貂脑病相似,Kurn病只见于新几内亚的高尔族人,尤其是女人,她们有吃人肉的风习,自从禁止了这种风习以后,此病的发病率明显地下降了。
2.研究自身免疫性疾病与遗传的关系
进行动物自发的自身免疫病的遗传学分析对于了解遗传因素在人类自身免疫病中的作用很有帮助。如(NZB×NZW)F1小鼠由于MHC决定的抑制性T细胞功能丧失或减退,能自发地发生与人的系统性红斑狼疮十分相似的自身免疫病,因此,一般认为它是人类自身免疫病的最佳天然模型。用系内同代互交和回交法可以推断:最低限度有三对等位基因控制免疫系统对自身抗原的反应;控制抗红细胞抗体生成的基因可能不同于控制抗核抗体生成的基因。这种推断也在一种莱克享有纯系鸡的自身免疫性甲状腺炎的遗传学分析中得到了证实。这种鸡的自身免疫性甲状腺炎的发病受Ir基因控制,它们缺乏控制性T细胞的功能。所以,B淋巴细胞能生成大量抗甲状腺自身抗体,通常这种鸡在孵出后二周即可查出抗甲状腺抗体,几个月后出现嗜睡、羽毛生长障碍,最后出现肥胖症及甲状腺机能低下的症状。如果在刚孵出时切除腔上囊,则可使这种甲状腺炎的发病率减低,或使其病情减轻,抗甲状腺抗体的效价下降。
3.研究胸腺功能低下与自身免疫病的关系
将新生小鼠摘除胸腺,造成T淋巴细胞缺乏的状态,可见小鼠逐渐地发生消耗病(Wasting Discase),其主要表现是体重下降,腹泻、反复发生感染。有些小鼠发生了与人的SLE相似的自身免疫性溶血性贫血、皮疹、肝炎及肾小球肾炎。但是,如果把去胸腺小鼠放在无菌环境中饲养,就不发生上述自身免疫性疾病。这说明对于自身免疫性疾病的发生来说,胸腺机能低落是重要的内在因素,而微生物的感染等则是重要的外界因素。又如,给新生的NZB小鼠摘除胸腺后,该小鼠的自发性自身免疫病加速发展,并迅速地出现溶血性贫血,血清中出现抗体DNA抗体、抗DNA组蛋白抗体、LE细胞及免疫复合物性肾小球肾炎。但是,如果把幼小无病的NZB小鼠的淋巴细胞输给这些小鼠,则可防止自身免疫病的发生,反之,如果病鼠的脾细胞(含T淋巴细胞在内)输给幼小无病的NZB小鼠,则小鼠将迅速持续地合成自身抗体。这些实验结果提示:胸腺功能衰退引起抑制性T淋巴细胞缺乏,与自身免疫病的发生之间有着密切的关系。与此相似,给刚刚孵出的莱克享肥鸡摘除胸腺时也能加重其日后出现的甲状腺功能低下。现在知道:胸腺功能正常是T淋巴细胞的正常分化、成熟所不可缺少的。如果胸腺功能减退,则胸腺激素的产生必然减少(事实上,SLE病人及NZB病鼠血中的胸腺激素水平是低的),故T淋巴细胞,包括抑制性T淋巴细胞的分化、成熟将发生障碍(停滞),结果,对自身抗原有反应能力的B淋巴细胞将不受遏制地产生大量自身抗体。
4.研究红斑狼疮疾病
对人的系统性红斑狼疮(Systgmic、Lupus Erythematous,SLE),长期以来一直使用有自发类似狼疮性肾炎疾病的B/WF1(NZB×NZWF1)小鼠作动物模型,研究病因和治疗方法,取得了很多资料。最近美国杰克逊实验室的Murphy医生,建立培育了有自发性红斑狼疮的MRL/MP/lprl(MRL/L)小鼠和BXSB小鼠(只有雄性鼠发生早期狼疮肾炎),用这些小鼠研究,得到了新的见解,重新评价了狼疮肾炎的发病机制。
现在至少有十几个品系的小鼠可以自发产生与SLE类似的自身免疫病,文献中报导的主要有四个品系,它们有各自不同的表现(见10-5、6)。表10-5中四种小鼠都是到一定年龄时发生SEL,表现为抗体上升,免疫复合物增多,肾炎,而后逐渐死亡,可是它们还有不同的发病特点,例如NZB小鼠主要还有较多的抗红细胞抗体,而MRL/L基本没有;MRL/L有类风湿因子、关节炎、淋巴组织增生,其它小鼠没有等。所以NZB可作为免疫性溶血性贫血的最好模型,MRL/L是研究类风湿的模型,NZB/W可以作为SLE的最好模型。此外值得注意的是胸腺对发病的影响也不同。幼小时切除胸腺,NZB/W的病有增加,说明切除胸腺后T细胞的抑制性降低,B细胞亢进,MRL/L去胸腺后不得病。
表10-5 四种SLE品系小鼠的表现
| 指标 | MRL/L | BXSB | NZB | NZB/W |
| 去 胸 腺 | 抗病 | - | 加重 | 加重 |
| 抗自身红细胞 | 0 | + | ++ + + | ++ |
| 类风湿因子 | + | 0 | 0 | 0 |
| 关 节 炎 | + | 0 | 0 | 0 |
| 淋巴组织增生 | 卅 | ++ | + | + |
| T、B细胞 | T↑ | B↑ | B、T↑ | BT、↓ |
表10-6 四种SLE品系小鼠自身混合淋巴细胞培养结果
| 小 鼠 品 系 | 年龄(月) | 转化细胞渗入3H-TdR(Pm×103) | |
| 自 发 | PHA刺激后 | ||
| C57BL/6 | 2 | 2.5 | 11.7 |
| CBA | 2 | 1.7 | 13.6 |
| NZB/W | 7 | 5.2 | 2.9 |
| MRL/L | 3 | 4.4 | 3.0 |
表10-6中四种小鼠的自身淋巴细胞混合培养反应性也不一样,说明它们的细胞本身有病变,不单有抑制性T细胞异常。
几种SLE品系小鼠的主要免疫学特性:
⑴NZB系:
来自N20第三代中的一对黑鼠。H-2d,淋巴细胞表面同种抗原(Thy-1.2,Ly-1.2,Ly2.2,Ly3.2,Qa-1a,Mlsa,Tlau),
①有自身免疫性溶血性贫血症:在4-5月龄以前Ht 值平均为44%,以后伴随增龄而下降,14月龄为32%,母鼠在6月龄后,公鼠在8月龄后出现Coombs氏试验阳性,伴随增龄阳性化率增高,10月龄为50%,12月龄以后达100%。自6月龄起出现抗核抗体。老龄小鼠出现红斑狼疮细胞。
②有高血清免疫球蛋白病:血清免疫球蛋白量异常之高,特别是IgM和IgG量递增。IgM量在一生中递增,其递增与性别、病情及Coombs氏试验都无关,并不因无菌饲养或初生期摘除胸腺而减少。血清中有抗2-和3-硝基酚抗体,且效价高。
③有类狼疮性肾炎:8月龄以后的小鼠自发与人的狼疮肾炎相似的以肾小病变为主的肾病变,并感染C型病毒。肾病变的为免疫复合物沉着所致。
④有特殊的胸腺病理组织学变化:伴随增龄胸腺重要较其它近交系小鼠为早期减小,髓质中形成淋巴滤泡,上皮细胞增殖,皮质中浆细胞和肥大细胞浸润。该现象与2~3周龄后出现的抗胸腺细胞自家抗体有关。
⑵NZW系
H-22,淋巴细胞表面同种抗原(Thy-1,2)。NZW与NZB杂交仔一代动物B/WF1(NZB×NZWF1),有自发的类似狼疮性肾炎、红斑狼疮细胞(LE细胞)阳性和抗核抗体阳性。
⑶BXSB系
来自C57BL/6J×SB/leF1,H-2b,淋巴细胞表面同种抗原(Thy-1,2、TL-、Ly-1,2、Ly-2,1,Ly-3,1、Qa-1b),抗自身红细胞阳性,淋巴组织增生,B淋巴细胞增多。
⑷MRL/1系
MRL/1小鼠是由C57BL/6J(H-2b)、C3H/Di(H-2k)、AKR/J(H-2k)及LG/J(H-2b)四个品系反复杂交育成的白化小鼠。从交配的过程推断,基因组成是LG75%,AKR12.6%,C3H12.1%,C57BL/60.3%。同种抗原和表现型是H-2k,Lyt-1.2+, Lyt-2.1+,Lyt-3.1+,Lyt-1.2+,TL-。IgG2a的同种异型为a。其它免疫学特性有:
①有全身性淋巴结显著肿胀:3月龄时肿胀明显可见,交随日龄增长而增大。
②有肾炎及血管炎、多呈急性或亚急性肾炎和多发性血管炎。4-5月龄时,用荧光标记可见抗IgG和C3抗体。在肾小球的肾毛细血管壁和肾小球膜上,能明显检出免疫复合物。血管内皮细胞和肾小球膜增殖,基底膜肥厚,有蛋白样物质沉着,类似人的狼疮肾炎。
③有类风湿关节炎:类似人的类风湿性关节炎,20-25%的MRL/1小鼠有关节软骨破坏,滑膜增厚,形成血管翳,渗出液贮留等类似人的类风湿关节炎的病状。
④有高免疫蛋白及补体减少:血液中免疫球蛋白量常呈高值、5月龄时达MRL/n小鼠的5倍,4~5月龄鼠的IgG达26mg/dl,约为正常小鼠的6-7倍。分类测定的结果是:IgA、IgM、IgG2b为正常小鼠的2倍,IgG1、IgG2a为6倍。血液中补体效价随月龄增长而下降。
⑤有半抗原抗体及自身抗体等。2-3月的MRL/1小鼠,在血清中检出抗2-硝基酚和抗3-硝基酚抗体。抗单链DNA抗体也在2-3月龄时被检出并随日龄而升。抗双链DNA抗体、抗Sm抗体;抗核糖核苷蛋白(RNP)抗体,抗ATP抗体等各种抗核成分的抗体效价也上升。
⑥有免疫复合物。血液中的免疫复合物可用多种方法检测到,MRL/1小鼠比其它狼疮小鼠的检出效价都高。免疫复合物与狼疮肾炎和血管炎都有直接关系。
⑦有淋巴细胞异常变化:MRL/L小鼠脾脏和淋巴结内Thy-1阳性的T淋巴细胞异常地增殖,4月龄以上小鼠的脾脏有70-90%,淋巴结有95%以上是Thy-2阳性细胞。自身免疫病小鼠的T细胞对刀豆素A的反应下降,而且用刀豆素A诱导,产生白细胞间介素Ⅱ的能力也下降。自发性狼疮肾炎小鼠的抗半抗原抗体,抗单链DNA抗体及产生免疫球蛋白的细胞增多特征为在初期是IgM型,而4月龄时则以IgG型为主,产生免疫球蛋白的细胞增多与肾炎的发展的有密切关系。
⑧免疫反应下降:对绵羊红细胞的一次及二次抗体反应答反应随月龄增长而显著下降。在2月龄时可查出对DNP--卵蛋白元的一次和二次IgG及IgE的应答反应,而到4月龄时就查不出的这些抗体了。
(5)MRL/n系
MRL品系小鼠在近交12代时发生常染色体隐性突变成两个亚系,其一具有lpr(lyromephoproliferative,淋巴增生)基因称MRL/1亚系,另一个则无此突变称MRL/n亚系。MRL/1小鼠因第5染色体上有lpr基因,2月龄时除发生全身性淋巴肿瘤外,并出现早期狼疮性肾炎,5-6月龄时半数死亡。疾病的发展比B/WF1小鼠快,所以使用价值也大。而MRL/n小鼠缺乏lpr基因,仅迟发轻度肾炎。Lpr基因为常染色体的劣性基因。把这基因导入其他小鼠,也能出现MRL/1小鼠的各种症状。
5.研究自身免疫病时的免疫病理特点
⑴多种自身免疫反应及其它免疫反应特点
共有的血清学特点为:高丙球蛋白血症、抗核抗体、抗dsDNA、抗SS-DNA、抗决定基抗体、高水平gp70、免疫复合物与补体水平降低(NZb C5缺乏)。
不同的特点为:出现抗Sm为MRL/n、MRL/1鼠系;出现IgG和IgMRA和关节炎的为MRL/1鼠系;出现抗红细胞抗体者为NZB、NZBXW鼠系;出现NTA者为NZB、NZBXW、BXSB鼠系,T细胞升高者为MRL/1鼠系,B细胞升高者为BXSB鼠系♀,B细胞及T细胞下降者为NZB鼠系♂、NZ鼠系。
⑵免疫复合物的致病作用
在小鼠SLE中,gp70-抗gp70的免疫复合物(IC)在伴随SLE的肾炎中起重要作用。在这类小鼠中15-30%具有急性或陈旧性心肌梗死或退化性血管病,此乃由于IC沉积于心肌血管所致。唯独MRL/1部分小鼠可发展为急性或坏死性关节性,甚至所出现的各种病变与类风湿关节炎十分类似。
⑶多株B细胞的高反应性
用三种方法测定出B细胞功能增高的现象;
①测定Ig分泌细胞及细胞(脾)质内Ig含量(免疫荧光法)细胞数量皆增高;②所有SLE鼠系脾脏抗三硝基苯(TNP)空斑形成细胞(PFC)皆含有较高水平的抗TNP结合SRBC的抗体;③鼠脾脏B细胞集落数目增多,B细胞的活跃尚表现于高丙球蛋白血症中;表现出对T-依赖性抗原去凝聚BGG及HGG的免疫耐受有抵抗力。更值得注意的是,MRI/1及雄性BXSB小鼠较正常MRL/n及雌性BXSB小鼠对免疫耐受性无抵抗力,容易诱导成功。有关全身B细胞活性增高和过度产生自身抗体的准确机制尚不清楚,但推测有可能是由于内源性或外源性B细胞激活、缺乏抑制T细胞负作用、增强辅助T细胞正反应、T调节细胞的亚类缺乏或缺乏免疫系统的其它成分如巨噬细胞等所致。但是,目前大多数认为B细胞的活性增高是与全身性T细胞的调节缺陷无关。
6.研究T及B细胞在自身免疫病的作用
通过实验动物模型研究证实自身耐受特别是T、B细胞的耐受(即对某一特定抗原无反应性)是引起自身免疫病发生的中心问题,虽然抑制细胞有时与耐受状态有关,但并非是诱导和保持耐受的原因。成年动物亦常会由应用无免疫原性抗原(可溶性或称去凝聚的蛋白质)引起耐受。诱发T、B细胞的耐受原的量具有很大不同,T细胞的耐受原量要比B细胞少得多,如小量抗原引起T细胞耐受时,B细胞仍旧活跃,并未诱导出耐受。例如在体内细胞外存在的低浓度自身抗原如甲状腺球蛋白、某些种类Ig或生长激素,皆可使T细胞产生耐受,而B细胞则不产生耐受,体内高浓度的血清白蛋白则可同时引起T、B细胞耐受。但是尚有一些存在于体内的很低浓度的抗原如碱性蛋白、细胞色素C、独特型决定簇等对T、B细胞皆不能引起耐受。总之T、B细胞的耐受程度是依赖于其微环境中自身抗原的浓度。
根据T、B细胞对自身抗原的免疫状态,实验性自身免疫疾病有以下结论:(1)实验性自身免疫甲状腺炎、EAT:T细胞耐受,B细胞激活;(2)实验性变态反应脑炎:直接激活活性T细胞;(3)实验性自身免疫重症肌无力、EAMG;T及B细胞皆激活;(4)类风湿性关节炎:单克隆B细胞激活。
(二)高<或低>免疫球蛋白血症
免疫球蛋白的变化对分析免疫反应极为重要,各种免疫蛋白(Ig)都具有一定的结构、特点及较稳定的数值,它是了解Ig各种免疫功能及免疫反应作用机制的物质基础。
人类和哺乳动物的血清中含有成千上万种免疫球蛋白,每一种免疫球蛋白分子的氨基酸排列顺序都不一样。按其理化、生物学及免疫特性可分为免疫球蛋白G、A、M、D、E五种。而IgG又可分为四个亚型,IgA分为三个亚型,IgM分为两个亚型。
小鼠免疫球蛋白中已知有IgM、IgA、IgE、IgG1、IgG2a、IgG2b和IgG3,其中IgE和IgG3正常小鼠血清中含量甚少。小鼠各类Ig的数量因品系不同而差异很大,因此了解用于实验的小鼠Ig的情况很重要。现将几个常用小鼠品系的各类Ig情况介绍如下:
1.IgM:在3月龄以前BALB/C、C3H/He、C57BL/6三品无差异,为200μg/ml左右。BALB/C在三个月以后也无多大变动。但C57BL/6随增龄而增加,生长一年雌性和雄性都超过500μg/ml,C3H/He伴随增龄也有一定程度增加,但其程度不如C57BL/6。此外,N2B呈高值,并出现IgM的巨球蛋白血症,CBA/N呈低值,出现IgG缺乏症。
2.IgA:BALB/C一月龄已呈高值,为C3H的2倍,C57BL/6的10倍。BALB/C、C3H/He和C57BL/6都随增龄而增加,尤以BALB/C为著,BALB/C在6月龄后,超过1000μg/ml的个体很多,8月龄全部超过1000μg/ml。皮下移植106IgA的浆细胞瘤细胞20天后,IgA量为2~3mg/ml左右,正常BALB/C超过1000μg/ml的个体可视为免疫球蛋白过多症。正常BALB/C最高值时(20月龄)也仅为200~300μg/ml,与BALB/C相比呈低值,C57BL/6伴随增龄缓慢增加,平均值不超过1000μg/ml。与雄性相比,BALB/C的雌性呈高值,其它品系无一定倾向。BALB/C的IgA呈高值,与BALB/C小鼠腹腔内注射矿物油后所生浆细胞瘤的60%为IgA合成株的现象有关。
3.IgG[XB]1[/XB]:C3H/He在3月龄以后下低,但BALB/C在3月龄以后仍然增加,和IgA相同,6个月龄以后几乎全部超过1000μg/ml。C3H/He在20月龄时为200μg/ml,降低到3月龄时1/5左右。C57BL/6为低值,但伴随增龄而缓慢增加。在IgG的亚型中,IgG1呈高值的品系有DBA/2,SJL和NZB。
SJL伴随网状细胞瘤而产生的单克隆性M成份,大部分为IgG,其中IgG1最多。在DBA/2×CBA杂交子一代,即使自然发生的浆细胞性白血病也几乎都是IgG,IgG1仍为最多。据报导IgG1难建立免疫耐受性,在SJL、BALB/C、NZB等品系难以建立免疫耐受性。
4.IgG2a:C3H/He6月龄以前呈高值。BALB/C在8月龄时呈短暂高值,在其它时期,看不到象IgA和IgG1那样与其它品系小鼠的显著差别。C57BL/6与C3H/He相比为低值,但随增龄而缓慢增加。
5.IgG[XB]2b[/XB]:BALB/C在8月龄时有一定增加,但在其它时间里几乎没有变动,为100~300μg/ml,在IgG的亚型中为最低值,C3H/He也看不出随龄而引起的变动。C57BL/6伴随增龄而增加,在IgG的亚种中为最高值,在12月龄以上的个体中有超过1000μg/mg者。
从上可知小鼠Ig含量多少因品系不同而差异很大。BALB/C的IgM和IgG2b为低值,IgG1和IgA为高值。然而C57BL/6则相反,IgG1为低值,IgG2b和IgM为高值。C3H/He的IgA为低值,IgG2a为高值。NZBIgM呈高值,出现IgM的巨球蛋白血症,CBAIgM呈低值,出现IgM缺乏症。
(三)各种免疫缺陷病和过敏症
1.补体(C′5)功能缺乏症:C5缺乏可使血清的调理作用失调,导致中性多核白细胞的趋化、吞噬及杀菌作用的无能,容易反复感染病菌。AKR/N和BIO、DZ/DSnN品系小鼠补体5缺损,易发生先天性补体C5功能缺乏症。DBA/2N品系小鼠补体C5活性低,相反BALB/CAnN品系小鼠的补体C5活性高。
2.迟发型超敏反应:迟发型超敏反应中包括结核菌素超敏反应,Jones-Mote型皮内过敏反应和接触性过敏症等。不同近交系小鼠的反应性有很大差异,如用纯蛋白衍生物(PPD)作抗原时,其足垫反应明显的近交系有:ICR,BALB/C,C57BL/6,DBA/2,C3H/He;反应稍弱的近交系为CFW,CDF1;反应弱的近交系为NZB,C57L,CBA;HR/Jms是反应最弱的近交系。如用绵羊红细胞(SRBC)作抗原时,不同近交系小鼠迟发型超敏反应也有较大差异,SWM/Ms、ddN、DDy是高反应的近交系;ICR、DDD、BALB/C(♀)为较高反应的近交系;C57BL/6J,C3H/He、DBA/2(♂)是低反应的近交系。
第五届国际免疫学会议重点研究讨论了速发型超敏感性的机理和调节。Sehon介绍了他的实验室用B6D2F1系小鼠做的工作。将变应原如卵青蛋白等与单丁氧聚乙二醇(mPEG)共价偶联后可抑制IgE的生成而不影响其他类抗体的水平。关于mPEG结合物用于治疗IgE介导的过敏性疾病的实用价值,已有作者在豕草和蜂毒过敏的病人中做过一些尝试,初步结果是令人鼓舞的。Sehon认为mPEG结合物不仅可用于Ⅰ型变态反应性疾病的治疗,而且可用于许多需注射异种蛋白质的疾病;如器官移植时为防止排斥作用而注射鼠或大鼠抗T细胞的单克隆抗体等。
K.Ishizaka介绍了他们实验室过去5年研究IgE应答的类特异性调节作用,在感觉巴西日园线虫的大鼠以及最近建立的能分泌IgE结合因子的大鼠-小鼠T细胞杂交瘤所作的工作。发现与IgE有亲和力的IgE-结合因子(IgE-BF)能选择性地调节IgE应答。他们将正常大鼠的肠系膜淋巴结细胞在体外培养时加入不同浓度的Cona (1r或10r/ml),经2-3天后取出此活化的细胞再加入IgE继续培养,发现两种浓度的ConA均能诱导产生IgE-BF,但IgE-BF的性质不同。低剂量Cona 所诱导产生的大部为IgE-抑制因子(IgE-SF),高剂量ConA诱导产生的大部为IgE-增强因子(IgE-PF)。前者不能与扁豆外源凝集素结合,能选择性地抑制IgE应答;后者能与扁豆外源凝集素结合并能增强IgE应答,其分子上有含甘露糖的低聚糖。作者进一步发现原来每ml加入10r的Cona IgE-PF,但在第二个阶段孵育时加入能抑制糖基化作用的蛋白质(Tunicamycin)后却改而产生IgE-SF。还证明福氏完全佐剂能促进产生IgE-SF,明矶能促进产生IgE-PF。这是由于前者能诱导产生糖基化作用抑制因子,而后者诱导产生糖基化作用促进因子。已知糖基化作用抑制因子系分子量为16000的多肽,可生成IgE-BF来调节IgE应答。Ishizaka希望对此课题的基础研究能开辟一条调节过敏病人IgE应答的新途径。
Katz过去证明在小鼠中有变态反应抑制因子(SFA)可抑制小鼠IgE的应答。在此次讨论中它以大量的实验证据证实了人SFA的存在。SFA能选择性地抑制IgE抗体合成。人SFA来自双向混合淋巴细胞培养(MLR)的上清液。测定系统是培养的人单个核细胞,于其中加入适量的PWM,在PWM刺激前一天加入MLR上清显著地抑制了淋巴细胞体外IgE的产生。Katz建立了一个敏感的放射免疫测定法,敏感度高达10pgIgE/ml。他的实验室已得到能产生大量人SFA的T细胞杂交瘤系,为研究人SFA的性质提供了方便,籍此采用遗传工程的方法将相应的转移到大肠杆菌,而生产大量的人SFA。
Kishimoto在已成功地建立了具有鼠IgE类特异性抑制功能的T细胞杂交瘤的基础上又建立了一些能分泌免疫调节因子的T细胞杂交瘤,并应用这个方法来克隆人IgE类特异性抑制性T细胞,初步结果表明已建立了能分泌IgE-抑制因子的T细胞杂交瘤。
Metzger进行了在IgF受体受到刺激后生物学方面的研究,发现一经改变了的抗体会使受体发生反应,而无须IgE的参与。
T.Ishizaka讨论了受体的搭桥及搭桥后膜止酶的活化。她的研究是鉴定与膜相关的那一个酶在Ca++涌入中所起的重要作用。用人的细胞进行此项工作受到细胞来源的限制。过去几年曾积极从事于建立人的嗜碱细胞和肥大细胞的体外培养,最近已能成功地培养人嗜碱细胞,使脐血的单个核细胞中的嗜碱细胞能选择性的生长。此项工作的要点是在全血培养中加入已除去IL-2的T细胞培养上清,经2-4周培养后有50-90%的细胞具有嗜碱细胞和肥大细胞的特点,但光学显微镜不能区分其为哪一种细胞,后经电镜观察证实为嗜碱细胞,因之称其为经过培养的嗜碱细胞。其表面标志为OKT3-、OKM1-、Thy1-,组胺含量为每100万细胞介于0.5-2.0r之间,每个细胞上约有12-38万IgE受体。此细胞与人IgE有高度亲和力,经人IgE致敏后用抗人IgE攻击可释放相当量的组胺。同时也发现此培养的嗜碱细胞不仅与人IgE结合,而且也可与小鼠和大鼠的单克隆IgE结合。小鼠IgE和人IgE结合到培养的人嗜碱细胞上的速率相似,只是小鼠IgE解离的速率较快。关于介质释放的启动机理仍有许多问题有待解决。但他相信有了大量有功能的可培养的嗜碱细胞,为进一步分析Ⅰ型变态反应性疾病所涉及的介质释放提供了一个有力工具。
Austen主要介绍最近几年对肥大细胞的新认识。已发现一个亚群的肥大细胞与一般含有肝素的结缔组织肥大细胞明显不同,它是受T细胞控制的。此第二类肥大细胞有产生白细胞三烯的独特能力,可能与粘膜上的肥大细胞是一致的。Austen将此类细胞称为E-肥大细胞,而将含肝素的称为H-肥大细胞。他发现,小鼠骨髓细胞在TH因子的控制下生成的肥大细胞在电镜下与一般的肥大细胞无差别。最近的研究是得知为维持此细胞或使此细胞分化增殖均需有IL-3(TH产生、分子量介于23000-29000的糖蛋白、等电点中性、活性很强、少量可使细胞分化),在小鼠骨髓细胞的培养中加入IL-3,于不同时间进行检查证实,8周后100%的细胞均具有IgE受体。
Austen还用同位素标记LTC4(白细胞三烯C4)检测证明,平滑肌上有LTC4的受体。由于了解到有第二类肥大细胞的存在以及对肥大细胞生物学的研究,使对速发型超敏反应的理解又前进一步。
3.X-连锁免疫缺陷:CBA/N小鼠有X-连锁免疫缺陷(X-linked Immune Defect),血清IgE水平很低,对某些抗原缺乏反应,如对胸腺信赖性(Thymus Dependent:TD)抗原的IgE和IgG缺乏应答性等。CBA/N小鼠这种X-连锁对多糖抗原反应缺陷具有重要意义,可以选用CBA/N来研究人的X-连锁缺陷病(如Bruton氏丙种球蛋白缺乏症、Wiskott-Aldrich氏综合症等)。因此CBA/N小鼠是研究X-染色体对免疫功能的影响和B细胞的发生、功能、异质性的极好动物模型。
4.过敏症:LEW大鼠对实验过敏性脑脊髓炎敏感,极易感染诱发性自家免疫性心肌炎,对诱发自家免疫性复合性肾小球肾炎敏感。易感染实验过敏性脑炎和药理诱发的关节炎。AS大鼠易感染实验过敏性脑脊髓炎,对自家免疫性肾小球炎敏感。AUG大鼠对实验过敏性脑脊髓炎易感,对自家免疫性甲状腺炎有抗力。WAG大鼠对实验过敏性脑脊髓炎有抗力,有些大鼠携带防御右旋糖苷过敏反应的隐性基因dx,对诱发自家免疫甲状腺炎敏感。
人类的一些自身免疫病的动物模型可参看表10-7。
表10-7 人类自身免疫性疾病的动物模型
| 疾 病 | 可能的抗原 | 动 物 模 型 |
| 自身免疫性溶血性贫血 特发性血小板减少性紫癜 | Rh、I、i、P 血小板膜 | 用李斯特单核细胞增多症菌免疫家兔、NZB小鼠、狗。 |
| 感染性脑脊髓炎 外周神经炎 | 中枢神经系统脑磷脂 外周神经髓磷脂 | 实验性变态反应性脑脊髓炎 实验性变应性神经炎 |
| 肾小球肾炎 | 肾小球基底膜,可能与链球菌抗原发生交叉反应 | 基底膜+福氏佐剂可引起羊肾炎、马杉肾炎 |
| 淋巴细胞性甲状腺炎 | 甲状腺球蛋白微粒体抗原,第2胶质抗原 | 肥鸡的甲状腺炎 变态反应性甲状腺炎 |
| 肝斗性肝硬化 溃疡性结肠炎 | 线粒体、胆管上皮细胞 结肠粘膜上皮细胞 | 用家兔抗线粒体抗体可造成大鼠的肝胆系统损伤。 用结肠上皮细胞或肠道上皮细胞免疫家兔及大鼠 |
| 交感性眼炎 水晶体葡萄膜炎 天疮疮 重症肌无力 部分男性不育症 | 葡萄膜抗原(色素?) 水晶体蛋白 皮肤的复层磷状上皮细胞间桥抗原 横纹肌 精子 | 同种葡萄膜组织加佐剂引起豚鼠发生本病。 在多种动物可造成变应性葡萄膜炎。 家兔模型 用肌肉抗原与2-硝酸复合物免疫家兔 实验性自身免疫性无精子生成症 |
(四)其他免疫性疾病
BUF大鼠:36周龄的雄鼠有自家免疫性甲状腺炎,大于一年龄的动物有26%发生自发性自家免疫性甲状腺炎和甲状腺单核细胞浸润。食入3-甲基五环碳氯化合物后能自发地发生自家免疫性甲状腺炎,而新生期胸腺切除后其发生率几乎达到100%,对绵羊红细胞缺乏免疫反应。
DA大鼠易患自家免疫性甲状腺炎。
PVG大鼠对诱发免疫性甲状腺炎有抗力。易感染溶组织阿米巴病。WAG大鼠对实验过敏性脑脊髓炎有抗力。有些动物可能携带防御右旋糖苷过敏反应的隐性基因dx,对诱发自家免疫性甲状腺炎敏感。对铁缺乏敏感,且多肽类(TG)-Pro-L继发性抗体反应性低。
五、在移植免疫研究中的应用
转植健康的器官以取代有严重的不可逆性病变而丧失功能的器官,是治疗疾病的一项主要措施。早在第二次世界大战期间对烧伤病人就进行了异体植皮,然而,这种移植全部以失败而告终。原因何在?1943年Medawar为了查明异体移植失败的原因,在家兔身上进行了一系列实验研究,明确了异体移植失败是因为受者对供者的组织发生了免疫反应。1953年Gorer及Shell首次断定:小鼠的异体移植失败,关键在于H-2抗原不相容。不同近交品系小鼠有不同的H-2型,二个相同H-2型品系小鼠间移植,可不发生排斥反应。
(一)移植的类型
根据供者(Donor)与受者(Recipient)的遗传学关系可以把移植分为四种类型:
1.自体移植(Autograft):为同一个体移植,如自体皮片移植。
2.同系移植(Isograft,Isogenic or Syngeneic Graft,Congeneic Graft):为同系异体间移植,如基因完全的一卵孪生子之间的移植,近交系内不同动物个体(基因型很相似的)之间的移植。
3.同种异移植(Allograft,Allogenic Graft,从前曾称为Honmograft):为同种异体间的移植,如同一种内的不同个体之间的移植,如鼠→鼠。
4.异种移植(Xenograft,Xenogeneic Graft,或Hetergraft),为不同个体间的移植,如不同种动物之间的移植,猩猩→人;猪→人等。
(二)动物对移植物的免疫排斥反应
受者的血管与供者的器官组织之间建立起血液循环之后,移植器官、组织的功能丧失,主要是由于免疫反应引起的损伤、坏死所致,其根据主要有以下几点:
1.给小鼠移植异系皮肤后,在头几天内受者的血管长入移植的皮片内,但从第3-4天起皮片内的血液灌流开始减少,皮片内的淋巴细胞及单核巨噬细胞浸润逐渐增多(浆细胞很少),并出现水肿、缺血、同时局部的引流淋巴结肿大,其内有大量淋巴母细胞的出现及核分裂。到了第9-10天以后,皮片发生坏死,脱落,这称为第一次排斥反应(First Set Rejection)。皮片脱落后,肉芽组织长入原来的移植部位,以后发生纤维化,形成疤痕;同时引流淋巴结也恢复原状。
2.如给该受者再次移植同一供者的皮肤,则在移植后的第3-4天提前出现排斥反应,而且强度比第一次强,血管很少或根本不长入移植的皮片内,皮片内很快地出现嗜中性白细胞、淋巴细胞及浆细胞浸润,并且血管内有血栓形成,这称为第二次排斥反应(Second Set Rejection,Secondary Boosting)。
3.如受者第二次接受另一供者的皮肤移植,则不出现第二次排斥反应,而是出现第一次排斥反应。
4.如给新生小鼠摘除胸腺,则等到该小鼠长大后接近异系皮肤移植时,不发生移植排斥反应;但是如果给去胸腺小鼠注射同基因型的正常小鼠淋巴细胞时,则仍能发生移植排斥反应,这表明淋巴细胞在移植排斥反应中起决定性的作用。
5.将已经发生过移植排斥反应小鼠的淋巴细胞注射到另一同基因型正常小鼠体内,然后再进行异系植皮时,则发生第二次排斥反应,皮片提早脱落,这提示:被异系皮肤致敏的淋巴细胞具有免疫回忆功能,即在第一次排斥反应中有一部分致敏淋巴细胞以回忆细胞的形式持续存在于小鼠体内。
6.在发生过移植排斥反应的小鼠血清中可查到针对供者组织相容性的抗原的特异性抗体,这种抗体能凝集供者的红细胞(在人,则对供者的淋巴细胞有细胞毒作用)。
上述实验结果无可辩驳地证明:移植排斥反应是一种特异性免疫反应。
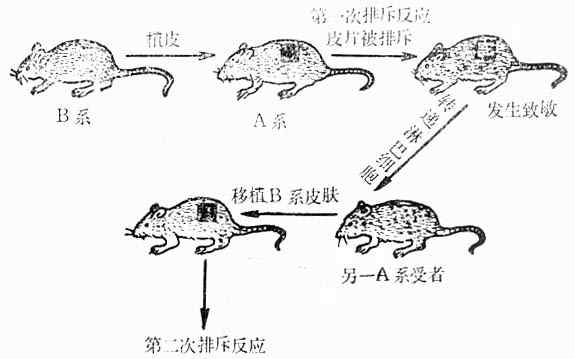
图10-4 第一次排斥反应及第二次排斥反应
(三)移植抗原(组织相容性抗原)
移植的异体组织之所以被排斥,是因为受者的免疫系统对供者的组织发生了免疫反应的结果。引起这种免疫反应的抗原称为移植抗原(Transplantation Antigens)或组织相容性抗原(Histocompatibctity Antigens)。在小鼠,这一抗原系统称为H-2抗原系统,一个H-2抗原分分由二个具有H-2特异性的肽和二分子β2微球蛋白构成。小鼠H-2抗原的分布密度以脾脏为最高,其次为肝、淋巴结、胸腺;再次,以肺、肾上腺、肾脏之顺序递减,红细胞、脑、睾丸、骨骼肌很少。
组织相容性(Histocompatibility)表示一动物种系各个成员的抗原个体性(Antigenic Individuatity),它决定于有核细胞表面的组织相容性抗原(移植抗原)的个体性。各种动物的主要组织相容性抗原都是由专门的基因决定的。决定主要组织相容的基因区称为主要组织相容性基因复合体(Majon Histocompatibility Complex,简称MHC)。小鼠的MHC座落在第17对常染色体短臂外端1/3,包括四个亚区。据估计,H-2基因复合体所含基因多达500余个,每一个区约含50~60个基因。H-2基因复合体的功能一是决定存在于细胞表面的同种异体抗原合成,另一是调控免疫反应(包括免疫反应及免疫抑制),二者之间是有关联的。因为很多免疫学特性是细胞膜功能的反应。
近交系和杂系小鼠异体移植的成败一般决定于下述规律:(参看表10-8和图10-5)
表10-8 接受组织移植的规律
| 供 体 | 受 体 | 接受或排斥 |
| 近交系(同性别) | 同一近交系 | 接受 |
| 近交系 | 其它近交系 | 排斥 |
| 近交系 | 本近交系与另一近交系交配生产的杂交一代(F1) | 接受 |
| 近交系??? | 同一近交系? | 接受 |
| 近交系?? | 同一近交系? | 排斥 |
| 杂交一代 | 杂交一代的亲本近交系 | 排斥 |
| 杂交二代 | 杂交一代 | 接受 |
| 杂交一代 | 杂交二代 | 排斥(少数接受) |
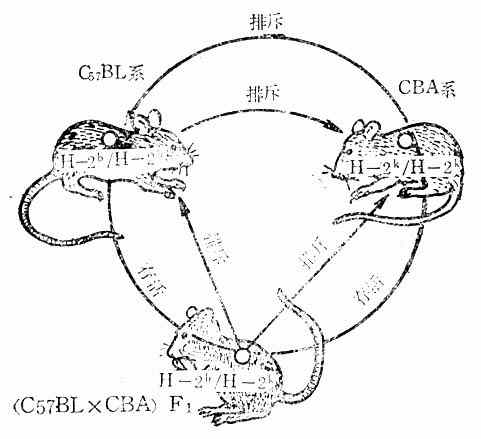
图10-5 不同近交系小鼠及其杂交一代之间的移植结果
⑴同一近交系的两个个体AA与AA(或BB与BB)之间进行器官移植时,移植物能够存活。
⑵不同近交系AA与BB之间进行移植时总是不成功。
⑶AA与BB的杂交第一代系F1(AB)与亲代之间进行移植时,如果AA→F1(AB)或BB→F1(AB),则因为供者有F1所含有的抗原成分,所以可以成功,但是,如果反过来,F1(AB)→AA或F1(AB)→BB,则不成功(图10-5)。
⑷F1(AB)之间进行交配所产生的F2,以及F3、F4……Fn,与F1(AB)之间进行器官移植时,可以成功。
⑸F1(AB)之间进行交配所产生的F2的基因型比例是AA:AB:BB=1:2:1。将F1(AB)与AA或BB进行回交时,其产生的下代基因型为AB:AA(或AA:BB)=1:1。将AA的皮肤移植给F2时,假定决定主要组织相容性抗原的基因位点只有一个,则F2中有3/4,回交子代中有1/2能接受AA的组织移植而不排斥。如果二个基因位点相同,则F2中有(3/4)2,回交子代中有(1/2)2能接受AA供者的组织而不排斥。如相同的基因位点为n个,则F[XB]2[/XB]接受AA移植物机率为(3/4)n,回交子代为(1/2)n。在小鼠,n=4~13(根据实验得出的)。这一事实意味着:在进行同种移植时,即使一定数目的组织相容性位点相同,而其它的位点不相同也有一定的成功机率。
当然,人类在遗传学上是复杂的,所以不能把上述法则推之于人类,不过,除了一卵孪生子以外,不能根据(5)得出移植必然失败的结论。如果部分组织相容性抗原相同的话,用免疫抑制药还有可能使异体移植获得成功。
(四)骨髓移植
骨髓组织中含有免疫活性的淋巴干细胞及淋巴细胞,所以,如果将异基因型的骨髓移植给免疫机能低下(包括原发和继发)的受者,则将对受者(宿主)的组织发生免疫排斥反应,结果引起GVH反应(Graft Versus Host Reaction,GVHR);如果给受者移植不含有免疫活性细胞的组织,则受者供者组织将发生通常所见到的那种免疫排斥反应,即宿主抗移植物反应(Host Versus Graft Restion,HVGR)。
在动物实验中可以造成GVH反应,它可以表现为矮小病(Runt disease)或消耗病(Wasting Disease)。矮小病是给初生动物(免疫功能还不够成熟)注射同种异系成年动物(免疫功能发育成熟)的脾细胞(相当一部分是下淋巴细胞),淋巴结细胞或胸腺细胞后而出现的,这时输入的T细胞与受者的移植抗原发生反应,结果,引起动物的发育不良。消耗病又称同种病(Homologous Disease),它可以通过先给成年动物进行X线全身照射以造成免疫机能低下的状态,然后输入同种异体的淋巴细胞而造成,此时动物出现营养不良、贫血、消瘦、恶病质,最后死亡。
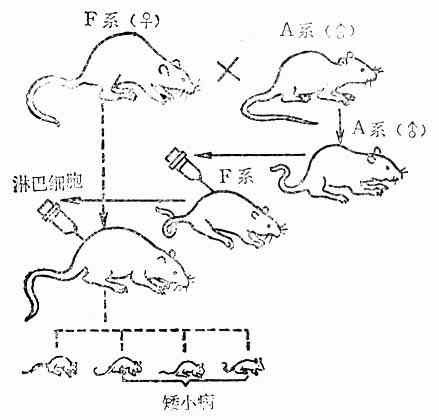
图10-6 大鼠的GVH反应产生矮小病的模式图